Life has exposed me to the tragic and devastating effects of Psychosis, both privately and professionally. During my time as a Support Worker, much first hand exposure to Psychosis has been part of the development path bestowed upon me.
These experiences lead to this topic and to seek White Paper evidence, which marries to the understanding which has been found. Which seems lacking by Healthcare Professionals generally, or ignored if understood.
Firstly, blood sugar, for several years, it seemed clear that severe Depression and Psychosis are Sugar fuelled, whether Hypoglycaemic driven depression, where the sugars are driven low for a prolonged period, or Hyperglycaemic where Sugars spike high, or worse spike and remain excessively high as seem in Psychosis.
The study below from 2002 is aptly linked as simply “Psychosis and Glucose” :
As Consultants in charge of Psychiatric Intensive care Units (PICU), we manage patients whose levels of arousal mean that they can not be managed on open acute psychiatric wards. All our patients have been compulsorily admitted under the terms of the Mental Health Act (England & Wales 1983).
We continuously monitor their physical condition, and this includes longitudinal monitoring of glucose levels. In addition, we monitor mental state using the Brief Psychiatric Rating Scale (BPRS), which gives an indication of the severity of the symptoms.
We report results in patients who have given their written informed consent to the publication of anonymised data, and who have agreed to form part of a study – approved by our local research ethics committee – of personal predictors on outcome in the PICU.
In forty-seven admissions to PICU, the glucose levels (mean 6.19 mmol/l SD 3.72)correlate with the BPRS on admission. If we exclude people with diagnosed non-insulin dependent diabetes, there is a direct correlation between BPRS at day 14 of the current admission and glucose levels during the first 7 days of the current admission (mean 5.13 mmol/l SD 0.93; p<_0.003 pearson=”pearson” correlation=”correlation” _0.477.=”_0.477.” p=”p”/>Twenty three admissions were treated with olanzapine (mean dose 32.85mg/day: BPRS 102.56 SD 29.13); twenty four were treated with risperidone (mean dose 7.58mg/day: BPRS 95.62 SD 14.29). At day 14, there were twenty one remaining (BPRS 72.38 SD 28.35) on olanzapine and twenty one on risperidone (BPRS 77.19 SD 20.09).
People with acute psychosis are highly aroused, and elevated glucose is not surprising. However, we suggest that control of glucose levels may be an important element of control of psychosis, and that elevated glucose levels – acute or chronic – may occur as part of the underlying psychosis, independently of the antipsychotic medication.
Psychotic patients often refuse to have blood tests. This can mean that, unless one is persistent in request for a sample (by a finger prick if necessary), one may miss hyperglycaemia.
Confronted with a psychotic patient who has abnormal glucose levels, psychiatrists may be tempted to modify or discontinue the antipsychotic drug for fear of precipitating a diabetic emergency.
We suggest that, on the contrary, adequate control of psychosis and disturbed behaviour with antipsychotic drugs may be necessary to prevent glucose spiralling out of control.
In addition, Convit et al (2003 PNAS 100: 2019-2022) have shown that reduced glucose tolerance may be associated with poor memory performance and hippocampal atrophy in elderly people who are not demented.
Hippocampal atrophy has been reported in psychotic disorders. Such studies do not, in the main, report the blood glucose, but it may be that, again as part of the underlying condition, disordered glucose may be pivotal.
Moreover, as hyperglycaemia is associated with both disturbed behaviour and microvascular disease, it is important to control for glucose in any assessment of the putative causal relationship of medication and microvascular (including cerebrovascular) disease.
/End Study
The first client supported with Psychosis, would regularly seem (to me) to have incidents, after meals with high Potato content and/or Sugar, having unprompted outbreaks of screaming and attacking anybody near.
Now, reflections of clients supported, also leads me to consider the second link, that of constipation, though more broadly Bowel disturbance in general, much like sugar instability, appears to aggravate behaviours, in certain clients the mental and behavioural impact is devastating.
Objective. Constipation is a prevalent problem in patients with psychiatric disorders; it reduces quality of life and may lead to severe complications.
The prevalence distribution of constipation across all psychiatric diagnoses in patients with severe mental illness (SMI) has hardly been studied. The aim of this study is to estimate the association between psychiatric disorders and constipation in SMI inpatients.
Methods. The strength of the association between constipation (based on use of laxatives) and DSM-IV psychiatric diagnosis was studied in a cross-sectional study with “adjustment disorders” as the reference group. The association was analyzed using logistic regression.
Results. Of the 4728 patients, 20.3% had constipation. In the stratum of patients older than 60 years, all psychiatric categories except for substance related disorders were significantly associated with a higher prevalence of constipation (odds ratios ranging from 3.38 to 6.52), whereas no significant associations were found in the stratum of patients between 18 and 60 years (odds ratios ranging from 1.00 to 2.03).
Conclusion. In the elderly, all measured psychiatric diagnoses are strongly associated with an increased prevalence of constipation. Physicians should be extra alert for constipation in SMI patients, independent of specific psychiatric diagnoses.
/End Study
Chooselife : It is my empirical experience that Bowel Movement(s) appear to have a huge impact on averting or avoiding incidents of Psychosis.
Constipation and dyspepsia are disturbing gastrointestinal symptoms that are often ignored in research on physical comorbidities of schizophrenia. The aim was to assess dyspepsia and constipation in a sample of outpatients with schizophrenia spectrum psychoses.
A general practitioner performed a thorough physical health check for 275 outpatients and diagnosed constipation and dyspepsia. This study assessed the possible contribution of several sociodemographic, lifestyle, and clinical variables to constipation and dyspepsia using logistic regression analysis. This study also assessed whether these symptoms were associated with abnormal laboratory findings.
The prevalence of constipation was 31.3%, and of dyspepsia 23.6%. Paracetamol (OR =3.07, 95% CI =1.34-7.02) and clozapine use (OR =5.48, 95% CI =2.75-10.90), older age (OR =1.04, 95% CI =1.01-1.06), and living in sheltered housing (OR =2.49, 95% CI =1.16-5.33) were risk factors for constipation. For dyspepsia the risk factors were female sex (OR =2.10, 95% CI =1.15-3.83), non-steroidal anti-inflammatory drugs (OR =2.47, 95% CI =1.13-5.39), and diabetes medication (OR =2.42, 95% CI =1.12-5.25).
Patients with dyspepsia had lower haemoglobin and haematocrit and higher glucose values than those without dyspepsia.
Patients with constipation had lower thrombocyte values than patients without constipation.
However, these findings were explained by factors pre-disposing to constipation and dyspepsia.
Clozapine use markedly increases the risk of constipation and may lead to life-threatening complications. In addition, analgesics and diabetes medication were related to gastrointestinal symptoms. These medications and their association to gastrointestinal symptoms should be kept in mind when treating patients with schizophrenia.
Schizophrenia is a serious long-term psychotic disorder marked by positive and negative symptoms, severe behavioral problems and cognitive function deficits. The cause of this disorder is not completely clear, but is suggested to be multifactorial, involving both inherited and environmental factors. Since human brain regulates all behaviour, studies have focused on identifying changes in neurobiology and biochemistry of brain in schizophrenia. Brain is the most lipid rich organ (approximately 50% of brain dry weight). Total brain lipids is constituted of more than 60% of phospholipids, in which docosahexaenoic acid (DHA, 22:6n-3) is the most abundant (more than 40%) polyunsaturated fatty acid (PUFA) in brain membrane phospholipids. Results from numerous studies have shown significant decreases of PUFAs, in particular, DHA in peripheral blood (plasma and erythrocyte membranes) as well as brain of schizophrenia patients at different developmental phases of the disorder. PUFA deficiency has been associated to psychotic symptoms and cognitive deficits in schizophrenia. These findings have led to a number of clinical trials examining whether dietary omega-3 fatty acid supplementation could improve the course of illness in patients with schizophrenia. Results are inconsistent. Some report beneficial whereas others show not effective. The discrepancy can be attributed to the heterogeneity of patient population.
Methods
In this review, results from recent experimental and clinical studies, which focus on illustrating the role of PUFAs in the development of schizophrenia were examined. The rationale why omega-3 supplementation was beneficial on symptoms (presented by subscales of the positive and negative symptom scale (PANSS), and cognitive functions in certain patients but not others was reviewed. The potential mechanisms underlying the beneficial effects were discussed.
Results
Omega-3 fatty acid supplementation reduced the conversion rate to psychosis and improved both positive and negative symptoms and global functions in adolescents at ultra-high risk for psychosis. Omega-3 fatty acid supplementation could also improve negative symptoms and global functions in the first-episode patients with schizophrenia, but improve mainly total or general PANSS subscales in chronic patients. Patients with low PUFA (particularly DHA) baseline in blood were more responsive to the omega-3 fatty acid intervention.
Conclusion
Omega-3 supplementation is more effective in reducing psychotic symptom severity in young adults or adolescents in the prodromal phase of schizophrenia who have low omega-3 baseline. Omega-3 supplementation was more effective in patients with low PUFA baseline. It suggests that patients with predefined lipid levels might benefit from lipid treatments, but more controlled clinical trials are warranted.
Polyunsaturated fatty acid (PUFA) metabolism abnormalities have been long implicated in the etiology of schizophrenia. Although several randomized clinical trials have been carried out to assess the efficacy of omega-3 PUFA as add-on therapy in reducing psychopathology in populations of chronic patients with schizophrenia, only a few concern first-episode schizophrenia. The majority of these studies used a 12-week intervention based on ethyl-eicosapentaenoic acid (ethyl-EPA), however, with conflicting results. An intervention based on docosahexaenoic acid plus EPA has not been used in first-episode schizophrenia studies so far. No add-on supplementation studies have been carried out in medicated first-episode schizophrenia patients to assess the efficacy of omega-3 PUFA in preventing relapses.
Methods
A randomized placebo-controlled one-center trial will be used to compare the efficacy of 26-week intervention, composed of either 1320 mg/day of EPA and 880 mg/day of DHA, or olive oil placebo with regard to symptom severity and relapse rate in first-episode schizophrenia patients. Eighty-two patients (aged 16–35) will be recruited for the study. Eligible patients will be randomly allocated to one of two intervention arms: an active arm or a placebo arm (olive oil). The primary outcome measure of the clinical evaluation is schizophrenia symptom severity measured by the Positive and Negative Syndrome Scale (PANSS). Other outcomes include depressive symptoms, patient functioning and the level of insight. Correlates of change measured during the study will include structural brain changes, oxidative stress and defense, as well as neuroplasticity indicators. Metabolic syndrome components will also be assessed throughout the study.
Discussion
By comparing 26-week administration of EPA + DHA or (placebo) olive oil as add-on therapy in reducing symptom severity and one-year relapse rate in patients with first episode schizophrenia, it is intended to provide new insights into the efficacy of omega-3 PUFA and correlates of change, and contribute to the improvement of mental health care for individuals suffering from schizophrenia.
Wenju Lu, Xiaoqing Liu, Tao Wang, Fei Liu, Airu Zhu, and Yongping Lin
Patients with coronavirus disease 2019 (COVID-19) exhibit a spectrum of respiratory symptoms like cough and dyspnea.1–3 Airway mucus is an adhesive viscoelastic gel composed mostly of high-molecular-weight mucous glycoproteins and water, which is important in maintaining lung function and health, pathological mucus hypersecretion may cause airway obstruction and lead to respiratory distress. Mucin (MUC) glycoproteins are the major macromolecular components of mucus, which are classified into two major types: the gel-forming secreted MUC5AC and the membrane-tethered MUC1.4 Here, with an attempt to understand the lung changes, we sought to provide a delineation of the components of airway mucus from patients with COVID-19.
To clean airway obstruction, respiratory tract mucus was aspirated and collected via bronchoscopy from COVID-19 patients with a critical illness, and optical coherence tomography (OCT) was applied via bronchoscopy to obtain cross-sectional images of the bronchiole. For healthy control, sputum was induced by inhaled hypertonic (3%) saline solution delivered with an ultrasonic nebulizer. After collection, sputum was processed as previously described for components analysis.5 Medical history, and clinical and laboratory data of the participants were extracted from electronic medical records. The study was approved by the Ethics Committee of the First Affiliated Hospital of Guangzhou Medical University (No. 2020-65). The requirement for informed consent was waived because the study was observational and the family members were in quarantine.
MUC5AC (sc-21701; Santa Cruz, Shanghai, China) and MUC1 (sc-6827; Santa Cruz) in airway mucus were measured using enzyme-linked immunosorbent assay, and MUC1-cytoplasmic tail (CT, clone EP1024Y; Abcam, Shanghai) levels were measured by Western blot analysis as described previously.5 The levels of MUC5AC and MUC1 were normalized to their average signal reading of a healthy control group.
A total of 16 patients with COVID-19 were included in this study, the clinical characteristics of the recruited subjects were shown in Table S1. There was no significant deviation in the distribution of age, or sex between the cohorts of case and control subjects. All the patients with COVID-19 were admitted to the intensive care unit because of low oxygenation index (199 ± 23 mm Hg), and 79% of them received mechanical ventilation. Blood laboratory tests showed elevated inflammatory indexes including leukocyte count, C-reactive protein, and interleukin-6 in most of the patients with COVID-19 (Table S2).
OCT indicated clear bronchiole in healthy controls (Figure 1A) and mucus retention in the bronchiole of patients with COVID-19 (Figure 1B). A volume of 1-8 mL white to gray sputum with high viscosity (Figure 1C) was aspirated from the respiratory tract of 16 patients with COVID-19. Induced sputum from healthy control was clear and transparent with low viscosity. Compared to healthy control, airway mucus from patients with COVID-19 had a higher level of MUC5AC (Figure 1D), MUC1 (Figure 1E), and MUC1-CT fragment (Figure 1F). However, there were no significant differences in the concentration of total protein, sodium, or chloride in the airway mucus from patients with COVID-19 when compared to healthy control (Table S3).
Representative cross-sectional images of bronchioles in (A) healthy and (B) critical ill COVID-19 patients were obtained with optical coherence tomography and bronchoscopy. C, Airway mucus aspirated by bronchoscopy. D, MUC1 and (E) MUC5AC protein levels as measured by ELISA, and (F) MUC1-CT level as measured by Western blot analysis in the airway mucus from COVID-19 patients and healthy control (HC). The levels of MUC5AC and MUC1 were normalized to their average signal reading of healthy control group. COVID-19, coronavirus disease 2019; CT, cytoplasmic tail; ELISA, enzyme-linked immunosorbent assay; MUC, mucin
Although more than half of patients with COVID-19 presented with a dry cough,6 this study provided direct evidence showing mucus retention in the small airway of patients with COVID-19, and patients were not able to expectorate by themselves and need bronchoscopy aspiration to help them to clean respiratory tract. The sputum from these patients with COVID-19 was viscous, which is not surprising as MUC5AC levels are extremely high, hyperconcentration of this gel-forming MUC dehydrates airway surfaces and causes mucus adhesion, which may contribute to airflow obstruction and respiratory distress. Clearance of airway mucus is an important way to increase oxygen and carbon dioxide exchange, bronchoscopy aspiration of airway mucus was used in all our patients to relieve hypoxia. In our center, all the 16 critical ill COVID-19 patients recovered and were discharged from hospitalization, which may attribute to our aggressive clearance of the respiratory tract. It is of note that bedside bronchoscopy may not be available in some hospitals as the medical resources are limited during the COVID-19 pandemic, carbocisteine has been reported to inhibit airway MUC5AC secretion, which could be used to reduce sputum viscosity and elasticity in patients with COVID-19. In addition, hydration of sputum by aerosolized hypertonic saline solutions or mannitol, and dilation of bronchi via aerosolized salbutamol may facilitate sputum expectoration.
MUC1 is a membrane-tethered MUC expressed on the apical surface of epithelial cells.7, 8 Since MUC1-CT fragment is on the cytoplasmic side of the cells, the elevated sputum CT fragment in patients with COVID-19 could come from detached and disrupted epithelial cells, which is evidenced by the pathological findings of diffuse alveolar damage with fibromyxoid exudates and macrophage infiltration in the lung tissue from patients with COVID-19.9
The limitation of this study is that induced sputum was used in the control group to compare airway mucus aspirated via bronchoscopy from patients with COVID-19, because it was very difficult for the patients with COVID-19 to expectorate sputum even with hypertonic saline solution inhalation.
The findings may suggest that increased level of MUCs in the airway mucus may contribute to the high viscosity of airway mucus and sputum retention in the small airway of patients with COVID-19, airway mucus clearance may be indicated to relieve respiratory distress, and MUC5AC may serve as a target for mucolytic agents in treating COVID-19. And MUC1-CT may serve as an indicator reflecting the severity of airway and alveolar epithelial cell damage.
ACKNOWLEDGMENTS
This study was supported by grants from the National Key R&D Project (2016YFC0903700 and 2016YFC1304102), the National Natural Science Foundation of China (81520108001 and 81770043), and grant specific for COVID-19 study from Guangzhou Institute of Respiratory Health. The authors would like to thank Dr Kwang Chul Kim (University of Arizona) for the invaluable assistance with the manuscript.
CONFLICT OF INTERESTS
The authors declare that there are no conflict of interests.
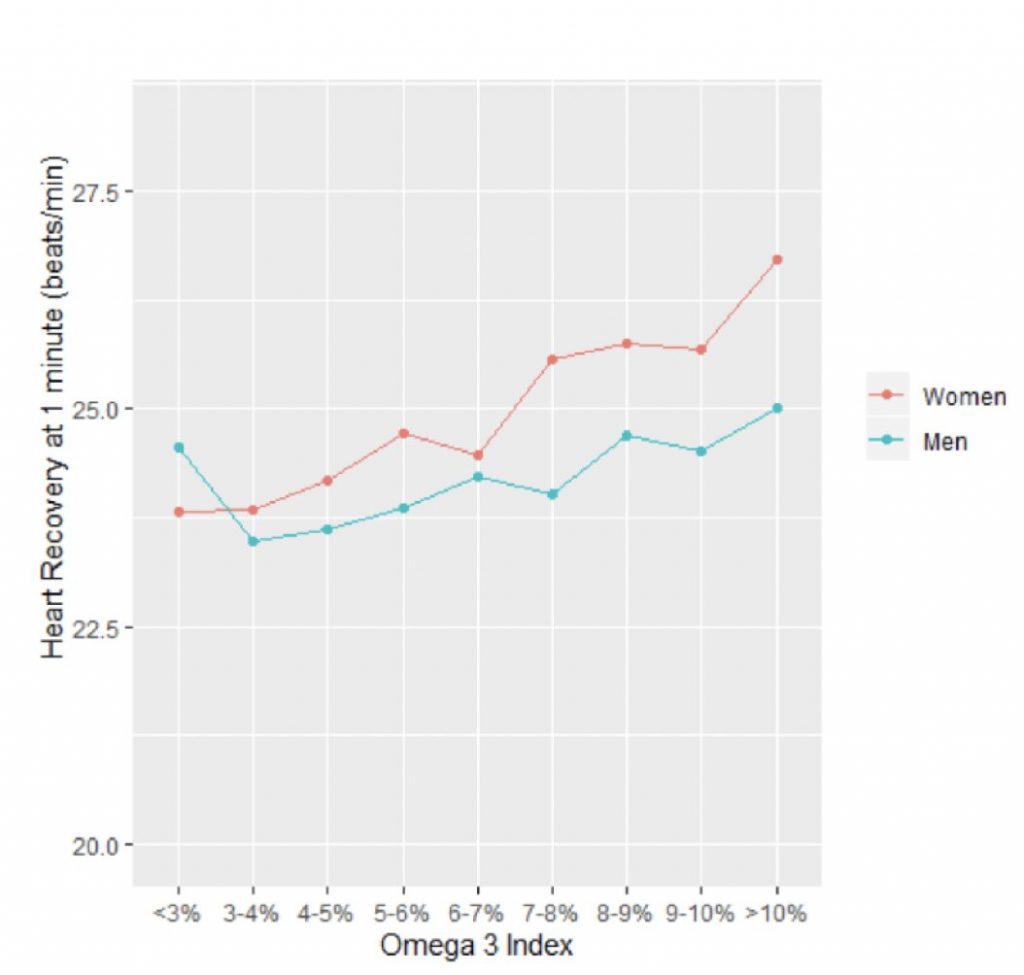
Comparing each patient’s heart rate recovery with their Omega-3 Index, the researchers found a significant relationship between these two variables such that the higher the Index, the faster the heart rate dropped down. The effect was more marked in women than men, but statistically significant in both. This is summarized in Figure 1 (right) where the Omega-3 Index is plotted on the x-axis, and the drop in heart rate one minute after stopping the treadmill test (measured in beats per minute, bpm) is shown as a positive number on the y-axis. Credit: Fatty Acid Research Institute
The Fatty Acid Research Institute (FARI) has published a new research paper in conjunction with The Cooper Institute on omega-3s and heart rate recovery.
Omega-3 fatty acids have a long history of being heart healthy, but exactly why and how has been less clear. They are known to lower serum triglyceride levels, but the effect is relatively small and it’s not clear how much of a risk factor high triglyceride levels is in the first place. So how do the omega-3 fatty acids EPA and DHA work?
A recent study from the Cooper Center Longitudinal Study (CCLS) and FARI sheds new light on this question.
The investigators utilized data from 13,912 healthy men and women who had preventive medical examinations at Cooper Clinic in Dallas over a 10-year period. These examinations routinely included both treadmill exercise testing and measurement of the Omega-3 Index (i.e., red blood cell EPA+DHA levels from OmegaQuant Analytics).
One component of the exercise test is called ‘heart rate recovery’ and it refers to how quickly the heartbeat slows down after maximal exercise. The faster it drops, the healthier the heart.
Comparing each patient’s heart rate recovery with their Omega-3 Index, the researchers found a significant relationship between these two variables such that the higher the Index, the faster the heart rate dropped down. The effect was more marked in women than men, but statistically significant in both. This is summarized in the Figure 1 (right) where the Omega-3 Index is plotted on the x-axis, and the drop in heart rate one minute after stopping the treadmill test (measured in beats per minute, bpm) is shown as a positive number on the y-axis. These are the unadjusted values. (For example, 25 bpm on the y-axis means that 1 minute after stopping exercising—i.e., the maximal heart rate—the person’s heart rate dropped by 25 bpm).
When adjusted for age, maximal METS (a measure of cardiorespiratory fitness), BMI and smoking status, a 2-percentage point higher Omega-3 Index was associated with a 0.35 and 0.69 bpm greater heart rate recovery in men and women, respectively (p<0.001 both).
Previous studies have shown that a slow heart rate recovery is associated with increased risk for sudden cardiac death, which fits with higher EPA and DHA levels being linked with reduced risk for sudden cardiac death3.
Dr. William Harris, President of FARI and co-inventor of the Omega-3 Index, who was also an author on the study, commented on the implications of this research. “These new findings from the CCLS harmonize with the known benefits of omega-3 fatty acids on resting heart rate and provide new clues to how these important fatty acids can preserve cardiac health,” he said.
“These benefits on cardiac autonomic tone join other cardioprotective effects of omega-3 fatty acids, including the reduction in blood pressure, chronic inflammation and platelet aggregation, to at least partially explain why omega-3s are good for the heart. Future treatment studies should define the omega-3 intake (and Omega-3 Index) that optimizes this aspect of cardiac function.”
Dr. James H O’Keefe, MD, a cardiologist and Medical Director of the Charles and Barbara Duboc Cardio Health & Wellness Center at Saint Luke’s Mid America Heart
Institute in Kansas City, MO, and a Professor of Medicine at the University of Missouri-Kansas City (who was not involved with the study) remarked that this research provides “strong data supporting the robust cardiovascular benefits of omega-3 fatty acids.”
The health effects of omega-3 fatty acids have been controversial. Here we report the results of a de novo pooled analysis conducted with data from 17 prospective cohort studies examining the associations between blood omega-3 fatty acid levels and risk for all-cause mortality. Over a median of 16 years of follow-up, 15,720 deaths occurred among 42,466 individuals. We found that, after multivariable adjustment for relevant risk factors, risk for death from all causes was significantly lower (by 15–18%, at least p < 0.003) in the highest vs the lowest quintile for circulating long chain (20–22 carbon) omega-3 fatty acids (eicosapentaenoic, docosapentaenoic, and docosahexaenoic acids). Similar relationships were seen for death from cardiovascular disease, cancer and other causes. No associations were seen with the 18-carbon omega-3, alpha-linolenic acid. These findings suggest that higher circulating levels of marine n-3 PUFA are associated with a lower risk of premature death.
Introduction
The n-3 polyunsaturated fatty acid (PUFA) family has been the subject of intense investigation ever since their inverse associations with risk for acute myocardial infarction were reported in Greenland Eskimos in the 1970s1,2. The PUFAs in this family include the 18-carbon, plant-derived alpha-linolenic acid (ALA,) as well as the 20–22-carbon, long-chain (LC, mostly seafood-derived) eicosapentaenoic (EPA), docosapentaenoic (DPA), and docosahexaenoic (DHA) acids.
The efficacy of the LC n-3 PUFAs in reducing risk for cardiovascular disease (CVD) remains controversial as findings from different randomized controlled trials (RCTs) have been conflicting. Nevertheless, a 2019 meta-analysis of RCTs reported significant reductions in risk for myocardial infarction, coronary heart disease (CHD) events and mortality, and CVD mortality in patients randomized to supplemental LC n-3 PUFAs3. Another meta-analysis of observational studies found that higher levels of circulating LC n-3 PUFA levels were significantly associated with a lower risk for CHD death4. However, no meta-analysis has yet examined the relationship between LC n-3 PUFAs blood levels and risk for all-cause mortality. Indeed, the only meta-analyses to report a beneficial association with all-cause mortality were based on the self-reported intake of fish5,6. Fish contain many nutrients besides just LC n-3 PUFAs, self-reported food intake is memory dependent, food databases can be out of date, and fish meals often replace less healthful choices. As a result, studies that link LC n-3 PUFAs and health outcomes based on self-reported fish intake have potential limitations. A more reliable and objective measure of LC n-3 PUFA consumption is their level in the blood7 which is primarily determined by the consumption of preformed LC n-3 PUFAs (although synthesis from dietary ALA can make a small contribution8). Hence a clearer picture of the biological relationship between LC n-3 PUFAs and disease outcomes may be obtained from biomarker-based investigations.
Some studies have reported inverse relations between n-3 PUFA biomarkers and total mortality9,10,11, while others have not12,13. In the Cardiovascular Health Study, higher LC n-3 PUFA levels also were associated with overall “healthier aging” (i.e., surviving past age 65 free of chronic diseases and maintaining good functional status)14. However, reports from studies of individual cohorts can be limited by insufficient power and inconsistent adjustment for potential confounding factors. In addition, publication bias can distort summary conclusions. To address these challenges, the present study pooled de novo individual-level analyses across 17 prospective cohort studies in the Fatty Acid and Outcome Research Consortium (FORCE)15 to explore the associations of circulating levels of n-3 PUFAs (both plant- and seafood-derived) and all-cause mortality. Secondarily, we examined the associations with mortality from CVD, cancer, and all other causes.
Here, we show significant inverse associations for all mortality endpoints with the LC n-3 PUFA levels. Hence, chronically higher tissue levels of these FAs operating through a variety of potential mechanisms may slow the aging process.
Results
Population
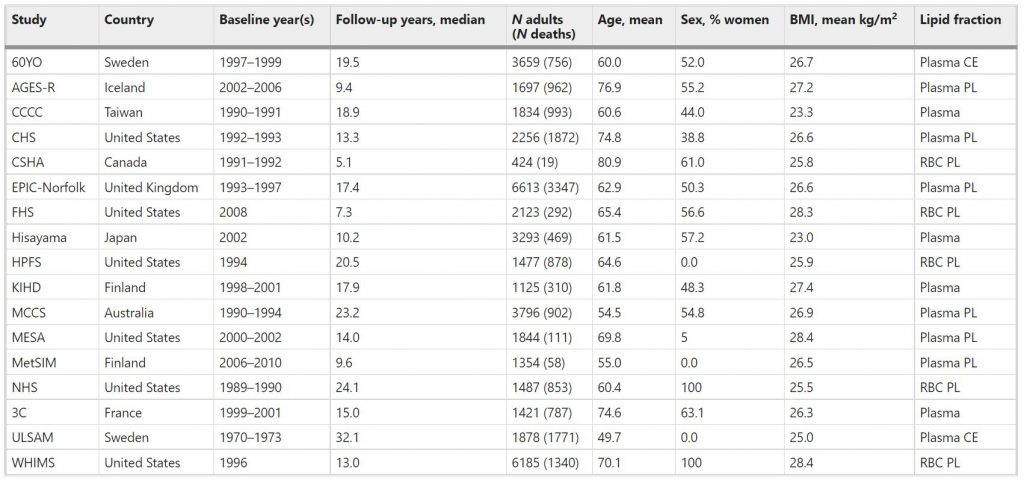
The pooled analyses included circulating n-3 PUFA measurements on 42,466 individuals, 15,720 (37%) of whom died during follow-up (Table 1). At baseline, the average age was 65 years (range of mean ages across cohorts was 50–81 years), 55% were women (range of 0–100% across cohorts) and the median follow-up time was 16 years (range of 5–32 years across cohorts). Whites constituted 87% of the sample. Circulating levels of the n-3 PUFAs (and of the n-6 PUFAs linoleic and arachidonic acids, which were included as covariates) are shown in Supplementary Fig. 1 and in Supplementary Table 2. Supplementary Table 3 shows the number of cause-specific deaths from participating cohorts. Overall, approximately 30% of the deaths were attributed to CVD, 30% to cancer, and the remaining 39% to all other causes.
Table 1 Baseline characteristicsa of 17 prospective cohort studies included in the meta-analysis: Fatty Acids and Outcomes Research Consortium.
aBaseline characteristics at the time of fatty acid biomarker measurement. Abbreviations of cohorts: 60YO, Stockholm cohort of 60-year olds, AGES-R Age, Genes, Environment Susceptibility Study (Reykjavik), CCCC Chin-Shan Community Cardiovascular Cohort Study, CHS Cardiovascular Health Study, CSHA Canadian Study of Health and Aging, EPIC-Norfolk European Prospective Investigation into Cancer, Norfolk UK, FHS Framingham Heart Study, HPFS Health Professionals Follow-up Study, KIHD Kuopio Ischemic Heart Disease Risk Factor Study, MCCS Melbourne Collaborative Cohort Study, MESA Multi-Ethnic Study of Atherosclerosis, MetSIM Metabolic Syndrome in Men Study, NHS Nurses’ Health Study, 3C Three-City Study, ULSAM Uppsala Longitudinal Study of Adult Men, WHIMS Women’s Health Initiative Memory Study. CE cholesteryl esters, PL phospholipids, RBC red blood cells.
Total mortality
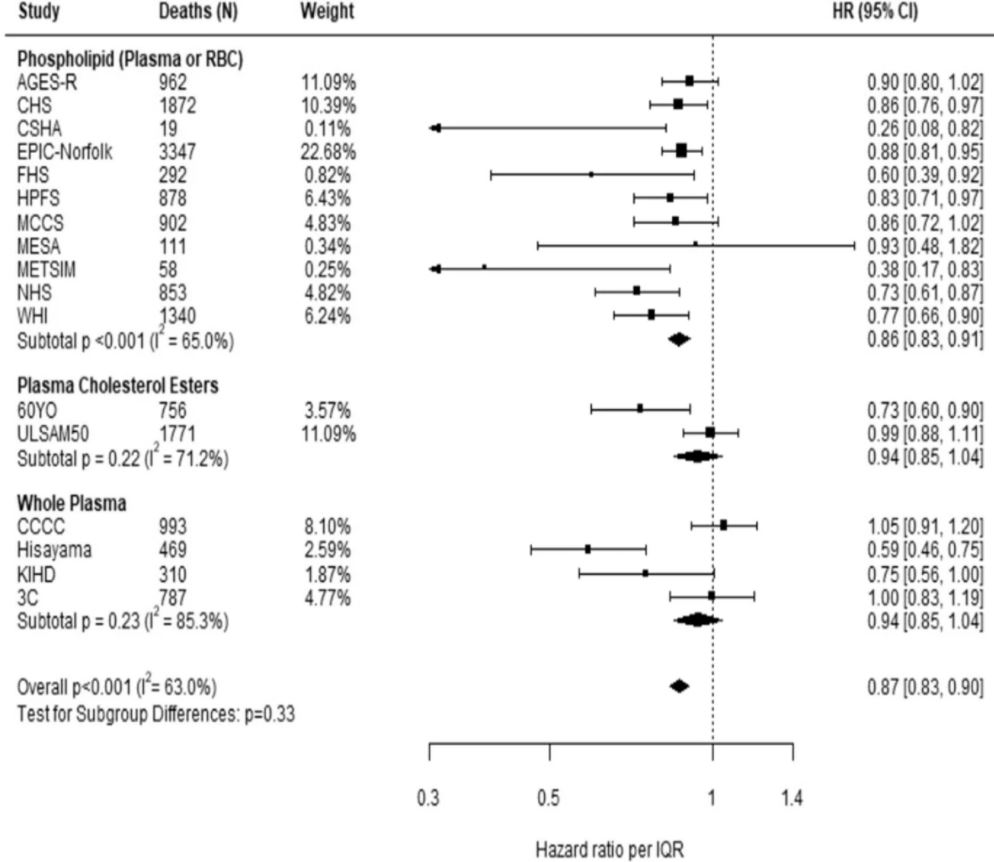
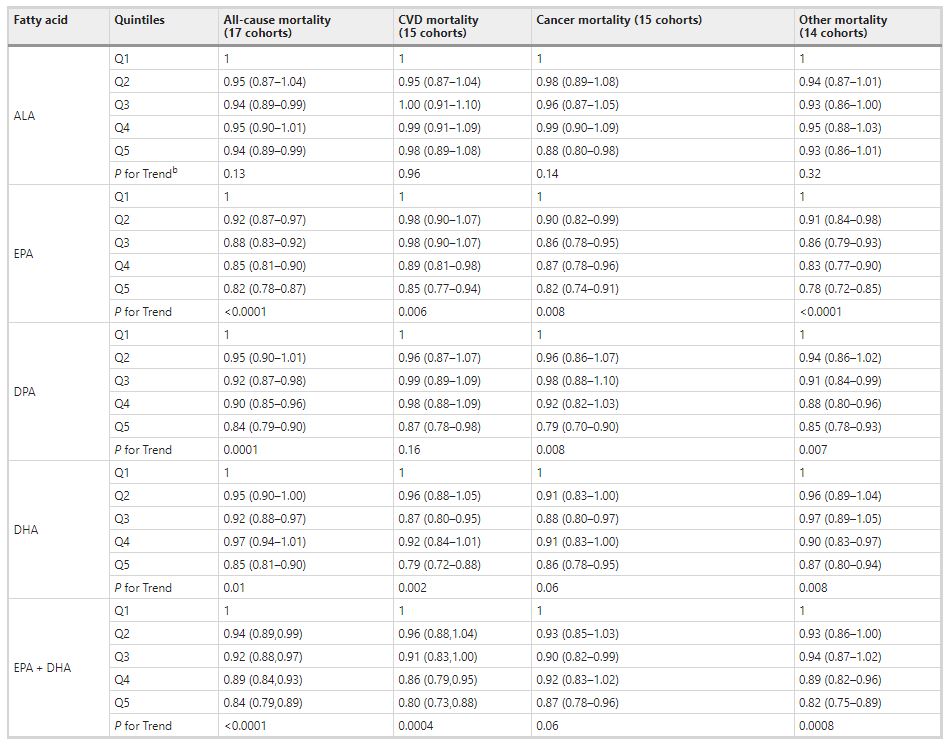
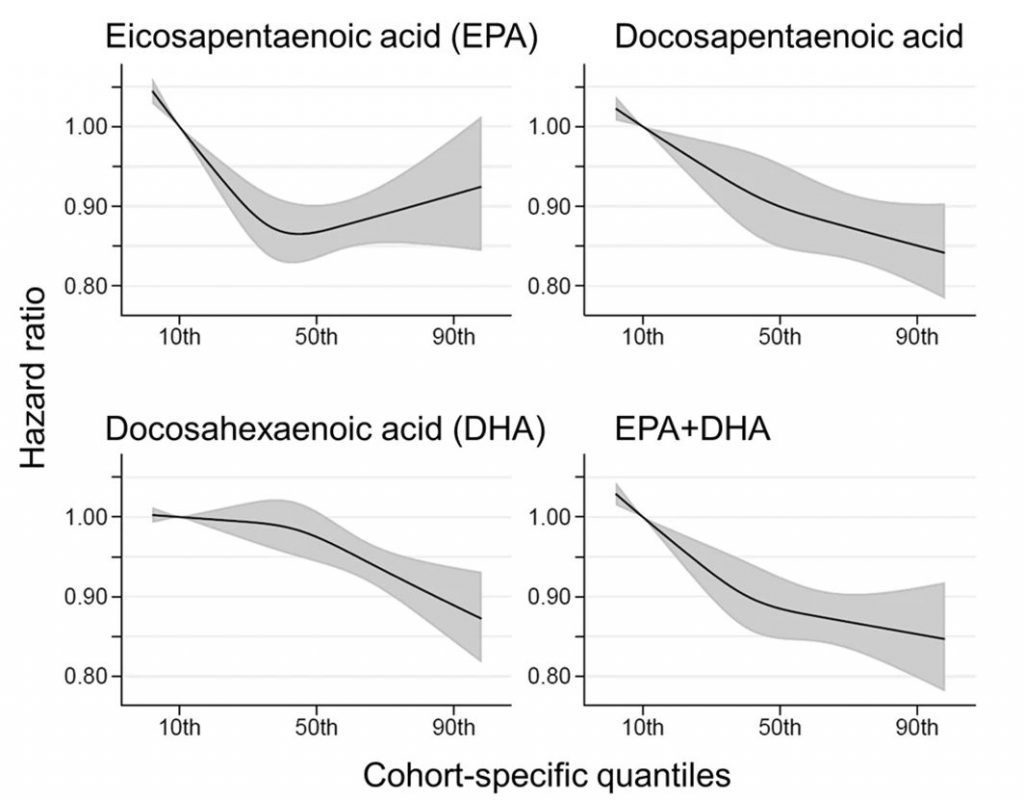
Comparing the medians of the first and fifth quintiles (i.e., approximately the 90th and the 10th percentiles), higher EPA, DPA, DHA, and EPA + DHA levels were associated with between 9% and 13% lower risk of all-cause mortality (Table 2). (The fatty acid levels associated with these percentiles for each cohort and sample type are shown in Supplementary Table 4). The HR for total mortality for EPA + DHA was 0.87 (95% CI: 0.83–0.90) (Fig. 1). In contrast, ALA was not significantly associated with all-cause mortality [HR 0.99 (0.96–1.02)]. In an across quintiles analysis, significant trends were observed for EPA, DPA, DHA, and EPA + DHA (all < 0.01); and comparing the top to the bottom quintile, each was associated with 15–18% lower risk of death (Table 3). There was little evidence for nonlinearity in these inverse associations for all each LC n-3 PUFAs except for EPA (p = 0.002 for the nonlinearity; Fig. 2). The relationship of EPA with mortality was most pronounced at lower levels and then appeared to plateau at higher levels. ALA was generally unassociated with total mortality, except for a borderline association in the top quintile [HR 0.94 (0.89–0.99); P-trend = 0.13], and there was no evidence for nonlinearity (Supplementary Fig. 2).
Hazard ratios (HRs) and 95% CIs expressed per cohort-specific inter-quintiles range comparing the midpoint of the top and bottom quintiles (see Supplementary Table 4 for cohort-specific n-3 PUFA values). All HRs are adjusted for age, sex, race, field center, body-mass index, education, occupation, marital status, smoking, physical activity, alcohol intake, prevalent diabetes, hypertension, and dyslipidemia, self-reported general health, and the sum of circulating n-6 PUFA (linoleic plus arachidonic acids). See Supplementary Table 4 for the 10th and 90th percentile values from each cohort for each PUFA of interest and the average PUFA values per lipid pool. Abbreviations: ALA alpha-linolenic acid, CI confidence interval, CVD cardiovascular disease, DHA docosahexaenoic acid, DPA docosapentaenoic acid, EPA eicosapentaenoic acid, HR hazard ratio.
Fig. 1: Adjusted hazard ratios (HR, 95% CI) for total mortality for circulating eicosapentaenoic (EPA) plus docosahexaenoic acid (DHA) in the 17 contributing studies of the Fatty Acids and Outcomes Research Consortium.
Study-specific estimates for HRs (dark squares) are shown per interquartile range (comparing the midpoint of the top to the bottom quintiles) their sizes indicate study weights (column 3). The horizontal line through each HR is 95% CI. Compartments included erythrocyte phospholipids, plasma phospholipids, cholesteryl esters, and total plasma. All HRs are adjusted for age, sex, race, field center, body-mass index, education, occupation, marital status, smoking, physical activity, alcohol intake, prevalent diabetes, hypertension, and dyslipidemia, self-reported general health, and the sum of circulating n-6 PUFA (linoleic plus arachidonic acids). See Table 1 footnote for abbreviations of cohorts.
Table 3 Meta-analysis of circulating n-3 PUFA biomarkers with mortality types by cohort-specific quintiles (hazard ratios and 95% CIsa): Fatty Acids and Outcomes Research Consortium.aExpressed per cohort-specific quintiles (see Supplementary Table 4 for cohort-specific n-3 PUFA values). All hazard ratios are adjusted for age, sex, race, field center, body-mass index, education, occupation, marital status, smoking, physical activity, alcohol intake, prevalent diabetes, hypertension, and dyslipidemia, self-reported general health, and the sum of circulating n-6 PUFA (linoleic plus arachidonic acids). bP-for trend is computed by using a fixed-effects, inverse weighted meta-regression analysis, i.e., the hazard estimates were regressed against study quintiles, which we assigned a value of 1, 2, 3, 4, or 5. Abbreviations: ALA alpha-linolenic acid, CI confidence interval, CVD cardiovascular disease, DHA docosahexaenoic acid, DPA docosapentaenoic acid, EPA eicosapentaenoic acid.
Fig. 2: Associations of circulating long-chain n-3 PUFA levels with all-cause mortality: nonlinear dose-response meta-analysis in the Fatty Acids and Outcomes Research Consortium. Hazard ratios and cohort-specific quantiles are presented in the vertical and horizontal axis, respectively.
The best estimates and their confidence intervals are presented as black lines and gray-shaded areas, respectively. The 10th percentile was selected as a reference level and the x-axis depicts 5th to 95th percentiles. Potential nonlinearity was identified for EPA (p = 0.0004) but not for the others (p > 0.05). All HRs are adjusted for age, sex, race, field center, body-mass index, education, occupation, marital status, smoking, physical activity, alcohol intake, prevalent diabetes, hypertension, and dyslipidemia, self-reported general health, and the sum of circulating n-6 PUFA (linoleic plus arachidonic acids).
Cause-specific mortality
Comparing the 90th to the 10th percentile, each of the LCn-3 PUFAs was significantly associated with a lower risk for death from CVD, cancer, and all other causes combined [except for DHA and cancer mortality, HR 0.93 (0.86–1.00)] (Table 2). ALA was not significantly associated with any cause-specific mortality. Evaluating the trend across quintiles, EPA, DHA, and EPA + DHA were inversely associated with CVD death, EPA and DPA were inversely associated with cancer death, and each of the LC n-3 PUFAs was inversely associated with other death. Comparing the top to the bottom quintile, EPA, DPA, DHA, and EPA + DHA were each significantly, inversely associated with CVD, cancer, and other mortality (Table 3).
Heterogeneity and sensitivity analyses
Inter-cohort heterogeneity was at least moderate (I2 > 50%) in the pooled analyses of all-cause mortality for all n-3 PUFAs except ALA (I2 = 26%) and EPA (I2 = 41%), while heterogeneity for cause-specific mortality ranged from little to moderate (0–56%) (Supplementary Table 5). There was little evidence of differential associations with mortality by PUFA lipid compartment after accounting for multiple testing (5 PUFAs × 4 outcomes; Bonferroni correction 0.05/20 = 0.0025, Supplementary Table 6). Likewise, associations of n-3 PUFAs with total mortality were similar across strata based on age, sex, race, and fish oil use (Supplementary Table 7), with no significant differences after accounting for multiple testing (5 PUFAs × 4 strata results; Bonferroni correction 0.05/20 = 0.0025). Overall findings did not change with the removal of participants taking fish oil (Supplementary Table 7) or in the drop-one-cohort analyses.
Discussion
In this meta-analysis utilizing a harmonized analytical strategy with individual-level data from 17 cohorts, we examined the associations between circulating levels of the n-3 PUFAs and mortality. We found that, after controlling for other major risk factors, LC n-3 PUFAs (but not ALA) were associated with about a 15–18% lower risk of total mortality comparing the top to the bottom quintiles. These relationships were generally linear for DPA, DHA, and EPA + DHA, but not for EPA. For this PUFA there was a steeper risk reduction across the lower blood levels but little additional difference in risk at higher blood levels. Inverse correlations were also generally observed between LC n-3 PUFA levels and CVD, cancer, and other causes of death.
This pooled analysis including over 40,000 participants and over 15,000 deaths greatly expands upon the findings of prior individual cohort studies that examined associations of circulating levels of n-3 PUFAs and all-cause mortality9,10,11,12,13,16,17,18,19,20,21,22,23,24. Relatively few studies have evaluated self-reported dietary fish (or estimated n-3 PUFA) intake in relation to total mortality, but those that have typically support our observations here5,22,25,26. Interestingly, reported use of fish oil supplements was linked to a lower risk for death from any cause in a study from the UK including over 427,000 individuals27.
Associations with total and cause-specific mortality were not significant for the plant-derived n-3 PUFA ALA. Prior biomarker-based meta-analyses reported inverse associations of ALA with CHD death, but relationships with total or CVD mortality were not examined4,28. Whether our finding of no association ALA on CVD mortality was because ALA has no role to play in fatal strokes (included in the CVD mortality metric) or because of differences in the cohorts included in these prior meta-analyses vs. the present one is not clear. Circulating ALA levels are less dependable markers of intake compared with the LC n-3 PUFAs because this fatty acid is rapidly β-oxidized and, to a small extent, converted into the LC n-3 PUFAs8. Nevertheless, the borderline and inconsistent relations of ALA on mortality risk deserve further study.
Higher circulating levels of LC n-3 PUFAs may beneficially affect diverse cellular systems that together could contribute to a reduced risk for death. The mechanisms behind the ostensibly beneficial effect of LC n-3 PUFAs on human biology are multiple and have been summarized in several recent reviews papers29,30,31,32. Among them are hypotriglyceridemic, antihypertensive, and antiplatelet effects; as well as positive effects on adipocyte biology, endothelial function, and autonomic balance. All of these appear to be mediated by effects on membrane physiochemistry, gene expression, and the production of a myriad of bioactive oxylipins. Persistently lower levels of inflammatory biomarkers also characterize those with higher circulating LC n-3 PUFA levels33. These fatty acids have been reported to inhibit the mammalian (or mechanistic) target of rapamycin (mTOR) in animal studies showing benefits in cancer34, metabolic syndrome35, spinal cord injury36, and depression37. mTOR inhibition extends lifespan in many species38 and acts as an energy sensor to coordinate gene expression, ribosome biogenesis, and mitochondrial metabolism39. In the Heart and Soul Study, where whole blood EPA + DHA levels were inversely associated with all-cause mortality24, higher levels were also linked with a slower rate of telomere shortening over a 5-year period40. As higher rates of telomere attrition have been associated with shorter overall lifespan41,42, this finding may be secondary to the more distal biochemical mechanisms noted above. Regardless of their specific actions, higher cellular levels of the LC n-3 PUFAs appear to slow the aging process.
Our findings of lower risk of CVD death with high vs. low blood levels of EPA + DHA are generally consistent with meta-analyses of self-reported fish intake25 and of biomarker levels4, as well as randomized controlled clinical trials of n-3 PUFA supplementation3,43 (although the most recent trial44 has not yet been included in meta-analyses). Compared with CVD, evidence for a link between n-3 PUFAs and cancer mortality risk is sparse, with no significant relationship for self-reported estimates of fish or n-3 PUFA consumption25,45. Meta-analyses of RCTs with n-3 PUFA supplements also have not observed effects on cancer, although short-term durations of such trials (generally up to 5 years) would likely preclude any ability to detect an effect on cancer46,47. The difference between these findings and what we observed may arise from the use of biomarker levels instead of self-reported fish intake. Biomarkers are potentially truer reflections of long-term exposure, making it easier to detect subtle relationships. In addition, circulating LC n-3 PUFA levels reflect endogenous metabolism, especially for DPA which is not correlated with estimated dietary DPA intake48 but may have important biologic effects49. Finally, since neurodegenerative diseases are a major non-CVD, non-cancer cause of death, a report that higher fish intake was associated with reduced mortality from this cause6 is consistent with our observations here.
Although circulating marine n-3 PUFA levels have not been measured in all of the major intervention trials, the doses of EPA + DHA used in most trials (<1 g/day) may not have resulted in marked differences in levels between treated and control patients50. For example, in the Vitamin D and Omega-3 Trial (VITAL) trial, treatment with 840 mg of EPA + DHA per day increased plasma phospholipid EPA + DHA levels from 2.7 to 4.1%, a 55% increase. This relatively small difference in LC n-3 PUFA levels between the placebo and active treatment groups could be one of the potential reasons for the failure of some RCTs to detect an effect of n-3 PUFAs on CV outcomes50,51. Future RCTs may be more effective if they focus on people with low baseline levels of LC n-3 PUFAs52 and provide doses of EPA and DHA that produce higher blood levels. An intake of about 250 mg of EPA + DHA per day as recommended in the Dietary Guidelines for Americans53 may raise circulating levels into the ranges observed here for some but not all adults7.
Although a significant effect on the primary (composite) endpoint in the VITAL trial47 was not achieved, our findings comport well with some of its secondary findings. In this study, the provision of 840 mg of EPA + DHA/day significantly reduced risk for major CV events and myocardial infarction in those participants with lower (vs. higher) intakes of fish (blood levels in these groups were not reported). There was a significant interaction of fish intake on total mortality as well; the HR (95% CI) in the low intake group was 0.87 (0.73–1.04) and in the high intake group, 1.19 (0.99–1.44, p for interaction 0.017). This secondary observation in VITAL implies that individuals with lower baseline LC n-3 PUFA levels are more likely to benefit from increased levels than those with higher baseline levels. Two recent RCTs examining the effects of high dose (~3–4 g/day) of LC n-3 PUFAs were performed in overweight patients with high blood triglyceride levels and at high risk for CVD events, all on background statin therapy. After 5 years of treatment, Bhatt et al.54 reported beneficial effects of EPA ethyl esters on CV events, whereas Nicholls et al.44 found no effect on the primary outcome using an EPA + DHA product in which the fatty acids were non-esterified. Another 2-year trial in elderly post-MI patients from Norway given 1.8 g of EPA + DHA found no benefit on CV outcomes55. None of these trials is directly relevant to our findings here owing to the nature of the high-risk patient populations, the number of concurrent background medications, the short duration of treatment, and the initiation of treatment late in life.
Strengths of the current analysis include the use of objective n-3 PUFA biomarkers (instead of estimated intakes from dietary questionnaires) which increases the accuracy of exposure assessment and allows for separate analysis of different individual n-3 PUFAs. The use of prespecified, harmonized, de novo individual-level analyses across multiple cohorts substantially increase generalizability, reduces confounding through consistent adjustment for covariates, and limits the potential for publication bias. The pooling of 17 studies including over 15,000 deaths also increased the statistical power to evaluate mortality subtypes as well as potential heterogeneity across subgroups.
Potential limitations deserve attention. Because our outcome was not rare, the hazard ratios (HRs) reported here (instantaneous relative risk) may be modestly different than the cumulative relative risk. Most individuals were White, potentially lowering generalizability to other races/ethnicities, although our analysis still included nearly 6000 non-Whites in whom findings for EPA + DHA were generally similar to those for Whites (Supplementary Table 7). Despite extensive efforts to harmonize study-specific methods, moderate heterogeneity remained between studies that may be due to unmeasured background population characteristics, differences in laboratory assessment of PUFAs and of outcomes, chance, or any combination of these. PUFAs and covariates were measured once at baseline, and changes over time could lead to misclassification, which could bias the results in uncertain directions. On the other hand, reasonable reproducibility has been reported for n-3 PUFA biomarker concentrations over time56. Because analytical methods, even within the same lipid fraction, were not standardized, and n-3 PUFA levels were measured in multiple fractions, we assessed cohort-specific n-3 PUFA percentiles rather than absolute percentages of total fatty acids in each fraction. Since FA levels were reported as a percent of total FAs in each lipid compartment, levels of one FA could affect levels of another. Indeed, in the plasma or RBC PL and CE pools, higher levels of the LC n-3 PUFAs (which were the focus of this study) are linked with lower levels of the n-6 PUFAs but not of saturated or mono-unsaturated FAs57,58. Since we adjusted for differences in linoleic and arachidonic levels in our analyses, this concern was accounted for. Each lipid pool used in this study reflects LC n-3 PUFA intake during relatively different and overlapping time periods generally from months to weeks following this hierarchy: RBC ≥ Plasma PL ≈ Plasma CE ≥ total plasma59,60. In addition, we cannot rule out the potential for residual confounding. That is, higher LC n-3 PUFA levels may simply be markers of a “healthy lifestyle,” and the fatty acids themselves may not be playing any physiological role in postponing death but would be biomarkers of a suite of other healthy behaviors (dietary/exercise/non-smoking, etc.), or endogenous metabolic processes, that might, in a multiplicity of ways, manifest in greater longevity. Although we adjusted for many major risk factors (age, income, marital status, smoking, hyperlipidemia, hypertension, etc.), residual confounding by other factors is always possible. However, the magnitude of the observed effect of the meta-analysis of circulating LC n-3 PUFAs and total mortality reported herein is consistent with the known associations with CHD mortality and sudden cardiac death61,62. Finally, as the attribution of cause of death is never as unambiguous as death itself, some uncertainty must attend to the cause-specific analyses reported here. In summary, in a global pooled analysis of prospective studies, LC n-3 PUFA levels were inversely associated with risk for death from all causes and from CVD, cancer, and other causes.
ChooseLife Notes : None of these studies, on ALA, will have included 37g odd emulsified 18-chain ALA daily, as Budwig studied and found to be the chief propellant to regain electron conductance within the cells. So whilst I like these studies they are biased towards Murine sources unjustly IMO, Flax has the highest level of surplus Electrons, or Pi Electrons, most enriching to our Electron Cloud, as Budwig variously described.
Donald Hathaway III, Krunal Pandav, Madhusudan Patel, Adrian Riva-Moscoso, Bishnu Mohan Singh, Aayushi Patel, Zar Chi Min, Sarabjot Singh-Makkar, Muhammad Khawar Sana, Rafael Sanchez-Dopazo, Rockeven Desir, Michael Maher Mourad Fahem, Susan Manella, Ivan Rodriguez, Alina Alvarez and Rafael Abreu
Corresponding Author: Donald Hathaway III, BSc. Division of Research & Academic Affairs, Larkin Health System, 7031 SW 62nd Avenue, South Miami, FL 33143, USA. Email: donald.hathawayiii@gmail.com
Infect Chemother. 2020 Dec
Abstract
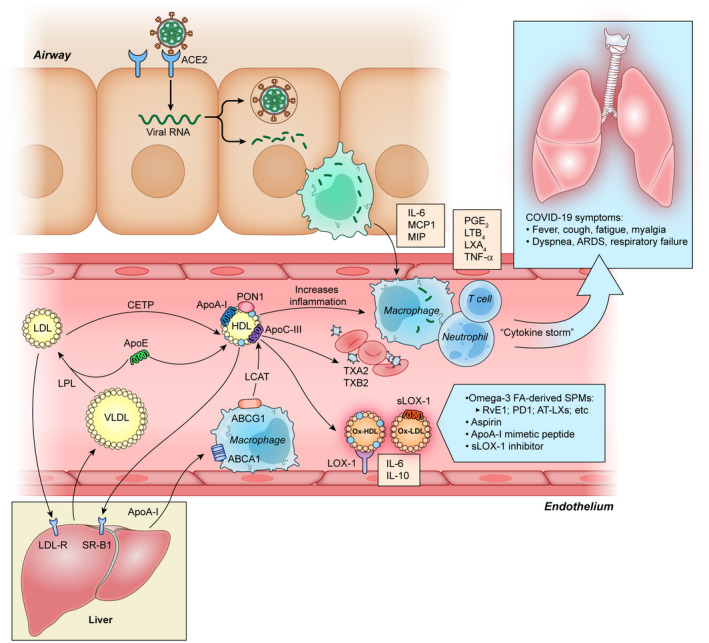
The rapid international spread of severe acute respiratory syndrome coronavirus 2 responsible for coronavirus disease 2019 (COVID-19) has posed a global health emergency in 2020. It has affected over 52 million people and led to over 1.29 million deaths worldwide, as of November 13th, 2020. Patients diagnosed with COVID-19 present with symptoms ranging from none to severe and include fever, shortness of breath, dry cough, anosmia, and gastrointestinal abnormalities. Severe complications are largely due to overdrive of the host immune system leading to “cytokine storm”. This results in disseminated intravascular coagulation, acute respiratory distress syndrome, multiple organ dysfunction syndrome, and death. Due to its highly infectious nature and concerning mortality rate, every effort has been focused on prevention and creating new medications or repurposing old treatment options to ameliorate the suffering of COVID-19 patients including the immune dysregulation. Omega-3 fatty acids are known to be incorporated throughout the body into the bi-phospholipid layer of the cell membrane leading to the production of less pro-inflammatory mediators compared to other fatty acids that are more prevalent in the Western diet. In this article, the benefits of omega-3 fatty acids, especially eicosapentaenoic acid and docosahexaenoic acid, including their anti-inflammatory, immunomodulating, and possible antiviral effects have been discussed.
Introduction
The coronavirus disease 2019 (COVID-19), now known the world over, is an emerging respiratory disease that was first identified in December 2019, in Wuhan, the capital of China’s Hubei province. It has since spread globally, resulting in the ongoing COVID-19 pandemic [1, 2]. In December 2019, this world-changing phenomenon began with an outbreak of pneumonia due to an unknown cause in Wuhan, with an epidemiological link to the Huanan Seafood Wholesale Market Place. The World Health Organization (WHO) was notified on December 31, 2019, by the Chinese Health Authorities [1]. The Chinese Center for Disease Control and Prevention identified a novel coronavirus on January 7, 2020, from the throat swab of a patient, which the WHO subsequently named 2019-nCoV [3]. This respiratory disease rapidly spread beyond the borders of China and by February 15, 2020, 26 countries were affected by this disease [4]. Considering this, the WHO declared it a public health emergency of international concern on January 30, 2020 and called for collaborative efforts of all countries to prevent the rapid spread of the virus [5]. Despite these efforts, the virus continued to spread, and the WHO declared it a pandemic on March 11, 2020 [2]. From its first outbreak in Wuhan, through November 13th, 2020, a total of over 52 million laboratory-confirmed cases of COVID-19 along with over 1.29 million associated deaths, have been reported globally [6]. In the United States (US), the total confirmed cases of COVID-19 have surpassed 10 million with over 240,000 deaths as of November 13th, 2020 [6].
The novel COVID-19 infection in humans may cause a wide range of symptoms, while some patients may remain asymptomatic even if they test positive for the virus. Possible clinical presentations may include fever, fatigue, sore throat, dry cough, shortness of breath, body aches, nasal congestion, anosmia, abdominal pain, and diarrhea, but other less common presentations have been reported as well. The minority of patients may however develop severe symptoms and serious complications such as interstitial pneumonia, septic shock, adult respiratory distress syndrome (ARDS), stroke, disseminated intravascular coagulation (DIC), and multi-organ failure (MODS). Strikingly, even asymptomatic patients are believed to be able to spread the disease [7].
Omega-3 fatty acids (FAs) are polyunsaturated fatty acids (PUFAs) that are abundantly available in nature. Omega-3 FAs belong to a category of supplements known as GRAS (generally recognized as safe). The Omega-3 Index Test serves as a measure of the amount of Eicosapentaenoic Acid (EPA) and Docosahexaenoic Acid (DHA) in the blood, especially in the red blood cell membranes. This index shows the ratio of omega-3 FAs to all other fatty acids. A recent study showed that more than 90% of American people consumed less than the recommended optimal value for omega-3 FAs in the diet (0.17 g/day) [8]. The metabolites of both omega-3 and omega-6 play an important role in the synthesis of different inflammatory mediators such as prostaglandins (PG), leukotrienes (LT), thromboxanes (TX), protectins, and resolvins. Omega-3 FA (fatty acid) plays a role in the host cellular membrane which regulates membrane fluidity and intricate lipid raft assembling in the cell membrane. A study conducted by Gutierrez and colleagues showed that omega-3 FA is incorporated throughout the body into the bi-phospholipid layer of the cell membrane of neutrophils and produces different mediators such as prostaglandins, leukotrienes, and maresins [9]. For that reason, if the injury occurs the byproducts of those cell membranes may produce less inflammatory provoking mediators compared to omega-6, which is more prevalent in the American diet [10]. Omega-3 FAs improve the function of the macrophages by secreting cytokines and chemokines, promoting the ability of phagocytosis, and activating macrophages by polarization [11]. Omega-3 FAs are also known to down-regulate Nuclear Factor-κ Beta (NF-κB). NF-κB is considered to be a transcription factor involved in cell signaling to initiate an inflammatory response by the innate immune system. The study shows that fish oil enhances antiviral response by inducing interferon (IFN) which inhibits viral replication [11]. Omega-3 FAs weaken the antiviral response of CD8 T cells and thereby could potentially be used to modulate cytokine responses to viral invaders [12].
A lack of omega-3 FAs in the diet can change the composition of the cell membrane. Every cell needs a healthy, functioning lipid bilayer to facilitate physiological responses and to maintain fluidity. However, the American diet contains high levels of omega-6 FAs as opposed to omega-3. One of the major causes of death in patients infected with severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) is multiorgan failure, which is a result of immune system overdrive causing cytokine storms. The omega-3 FA is known to produce less pro-inflammatory cytokines, therefore increasing omega-3 FA intake in the diet or supplementation could decrease viral entry, promote better immune function, and decrease severity among those who have been diagnosed with COVID-19. As we are still searching for definitive treatment, omega-3 FAs might be a safe and relatively inexpensive prophylactic and treatment approach for those who are at high risk and those who have the disease. This review aims at describing the health benefits of consuming a diet rich in omega-3 FAs in addition to the possible role in COVID-19.
Coronavirus in Humans
Coronaviruses are a family of spherical viruses, and their surfaces are covered with ‘crown-like’ spikes. These viruses are enveloped with single-stranded linear positive-sense RNA genomes. They are classified under the order Nidovirales, family Coronaviridae, and subfamily Orthocoronavirinae [13]. They have the largest genome for RNA viruses and based on genetic and antigenic criteria, they have been divided into four genera: alpha, beta, gamma, and delta Coronaviruses [14]. They have been widely distributed in nature and can affect other species like birds, bats, cats, rodents, pigs, and other mammals [15]. The seven coronaviruses that infect humans are SARS-CoV-2, alphacoronavirus 229E and NL63, and betacoronavirus HKU1, OC43, coronavirus associated with the SARS-CoV and coronavirus associated with respiratory syndrome in the Middle East (MERS -CoV) [16].
A wide range of similarities exists between SARS-CoV and SARS-CoV-2, as shown in Table 1. Both of them belong to the family Coronaviridae [17]. Studies show that there is a 76.47% similarity in amino acid sequences in both of them [18]. Comparison of RNA sequence and protein modeling showed that the domain on the spike protein which is responsible for attaching SARS-CoV-2 to the receptors on the host cell is similar to SARS-CoV [19]. Both use angiotensin-converting enzyme 2 (ACE-2) receptors for entry in the human respiratory epithelium cells [5, 17]. They are transmitted by respiratory droplets and contact with infected people [20, 21] and responsible for causing symptoms like fever, cough, shortness of breath, and fatigue [22]. They cause the appearance of ground-glass opacities on the Chest X-ray. The simple way to prevent them is by hand hygiene, social distancing, and respiratory hygiene [21]. MERS-CoV uses Dipeptidyl peptidase-4 (DPP4), CD26 receptors to attach to the respiratory epithelial cells, and pneumocytes [17].
SARS-CoV, severe acute respiratory syndrome coronavirus; MERS-CoV, Middle East respiratory syndrome coronavirus; SARS-CoV-2, severe acute respiratory syndrome coronavirus-2; N/A, not applicable; ACE-2, angiotensin-converting enzyme 2; DPP4, dipeptidyl peptidase 4; CD26, cluster of differentiation 26.
The basic reproduction number (R0) of COVID-19 is between 1.4 to 2.5 according to the statement based on January 23rd, 2020. R0 is much higher for COVID-19 compared to that of SARS (1.7 – 1.0) and that of MERS (<1). COVID-19 has been a clinical mystery, as of now, with unique epidemiology, pathogenesis, and clinical outcomes [23]. This type of attachment enhances viral fusion to the human cells and atypical severe clinical outcomes in the host. ACE-2 receptors are present in the heart, lungs, gastrointestinal tract, and blood vessels. ACE-2 receptors are the receptors that mediate the viral entry of SARS-CoV-2, causing vasoconstriction, inflammation, and thrombosis [17, 24].
The diameter of the virus is 125 nm. The 3-D structure shows that nucleocapsid protein and nucleic acid are found beneath lipid bilayer [25]. It contains 14 functional open reading frames (ORFs), out of which two ORFs make replicase genes responsible for encoding proteins needed for the synthesis of viral RNA. The other 12 ORFs are responsible for making eight accessory proteins and four structural proteins: membrane, spike, envelope, and nucleocapsid [26, 27]. One of the main virulence factors of coronavirus is N protein, which is highly infectious [28]. The structure of the coronavirus spike is very complex, with three main segments. These three segments consist of a short intracellular tail, single-pass transmembrane anchor, and a large ectodomain. The ectodomain contains a receptor-binding subunit S1 and a membrane-fusion subunit S2. The coronavirus spike on electron microscopy is a clover-shaped trimer with three S1 sections and a trimer. When the coronavirus infects a host cell, it loosely binds to the receptor via the S1 subunit and the S2 subunit connects the ACE-2 receptors on the host cell with the coronavirus cell membrane. This allows for the integration of the coronavirus genome with the host cell genome [19, 29, 30]. ACE2 receptors are present in the nose, lungs, blood vessels, intestines, and certain areas of the brain [31]. It uses Transmembrane protease serine 2 (TMPRSS2) serine protease to prime S protein [32].
Accumulating evidence suggests that SARS-CoV-2 is most likely a zoonotic source from the wet market in Wuhan. A vast number of people were exposed to this animal marketplace. This proposes the idea of the animal to human transmission at some point likely being the primary source of spread [33]. According to Rothan and Byrareddy, SARS-CoV-2 is primarily transmitted via person-to-person direct contact through respiratory droplets by cough and sneezing [33]. Numerous case studies have also indicated the presence of SARS-CoV-2 live viral RNA in feces. This is suggestive that there is a high probability of fecal-oral transmission with SARS-CoV-2 as another possible route of transmission [34]. The novel Coronavirus is a developing situation where through data analysis and time we will be able to understand more possible routes of transmission. According to Qu and colleagues, strong evidence suggests that the coronavirus can thrive for extended lengths of time outside of its host cell. It is also believed that the COVID-19 virus can survive for many hours on a large number of surfaces including sterile sponges, aluminum, or latex materials. This increases the virus’ opportunity for transmission from the external environment into the host cell through contact with the eyes, mouth, and nose [35]. A study by van Doremalen and team analyzed surface integrity and aerosol of SARS-CoV-2 compared to SARS-CoV. This study utilized Bayesian regression to measure the decay rates of both viruses in aerosol and surfaces. COVID-19 showed viability in aerosol for up to 3 hours, with a decay rate of 103.5 to 102.7 Median Tissue Culture Infectious Dose (TCID)50/L . This decay rate was very similar to the decay rate seen in SARS-CoV, which was 104.3 to 103.5 TCID50/mL suggesting that aerosols can remain infectious for several hours and surfaces can remain infectious for up to one day [36].
Every person is at risk to be infected with SARS-CoV-2 if exposed; however, not everyone develops severe, life-threatening infections [37]. Patients who are more susceptible to develop severe infections include patients with comorbidities like diabetes mellitus, chronic obstructive pulmonary disease, interstitial lung disease, chronic heart failure, coronary artery diseases, cardiomyopathies, metabolic syndrome, and hypertension [37]. The older age group (especially above 85 years of age) and with underlying medical conditions like chronic kidney disease, sickle cell disease, cystic fibrosis, cerebrovascular diseases, liver diseases, smoking, thalassemia, neurologic diseases like dementia and immunocompromised state due to transplantation of solid organ, obesity (body mass index 20 or greater), use of steroids/other immunomodulatory drugs, human immunodeficiency virus, blood or bone marrow transplant are also at increased risk [6, 37, 38]. The SARS-CoV-2 life cycle into host cell is shown in Figure 1.
Figure 1. Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) spike (S) glycoprotein binds with host cell angiotensin converting enzyme 2 (ACE-2) receptor. Subsequently, the virus enters the host cell either through membrane fusion or endocytosis and releases its positive sense ribonucleic acid (RNA) in the host cell cytoplasm via uncoating. The host ribosomes translate viral polymerase protein from positive sense RNA. The viral polymerase replicates negative sense RNA from the positive sense RNA. The viral polymerase then utilizes negative sense genome to produce more positive sense RNA (genomic replication) and mRNAs for nucleocapsid (N), spike (S), membrane (M), envelope (E) (subgenomic transcription). After the translation of viral structural proteins, S, E, and M proteins are processed in Endoplasmic Reticulum-Golgi (ERG) intermediate compartment. Nucleocapsids are assembled in the cytoplasm and then bud into the lumen of the ERG intermediate compartment. Finally, the mature virus inside the Golgi vesicle is released from the infected cell through exocytosis. A mature virus is capable of infecting the lung, endothelium, intestine, heart, testis, kidney through ACE-2 receptors.
Omega-3 Fatty Acids
Omega-3 FAs might be a safe and relatively inexpensive prophylactic approach for those who are at high risk. For a problem that has arisen from nature, we may return to nature for the cure.
Omega-3 FAs have been investigated repeatedly since 1994 when the tremendous health benefits were first established. They were found to lower the risk of thrombosis in cardiovascular disease and it may impact positively on inflammatory diseases, brain function, and mental health, among a multitude of other benefits [39]. The term omega-3 comes from the structural descriptor for a family of PUFAs. Within the omega-3 FA family, we can find and linolenic acid (LNA) and its derivatives, including α-linolenic acid (ALA), eicosapentaenoic acid (EPA), and docosahexaenoic acid (DHA). All of which are crucial elements for the functions of various cells and organs such as the brain, eyes, cardiovascular system, immune system, and general human growth [40]. In that sense, it has been shown that neural membranes of our neurons contain high concentrations of PUFAs. The omega-3 FAs belong to the essential fatty acid group within the PUFAs family and previous studies have demonstrated that our bodies cannot synthesize them. Therefore, they must be obtained from the diet [41].
Extensive evidence has been published about levels of omega-3 PUFAs that can mediate anti-inflammatory effects [9, 40, 41, 42, 43]. For example, omega-3 FAs play a role in mediating inflammatory processes and immunomodulation for both innate and acquired immune systems [9]. Also, a study done by Chanda and team concluded that omega-3 FAs could be a potential antimicrobial drug with little potential for drug resistance [44]. However, the use of these fatty acids as antimicrobial agents has not yet received much attention in clinical medicine worldwide.
Nowadays, the novel coronavirus has resulted in a pandemic that has spread with sheer rapidity across the globe. In this literature review, we provide information on omega-3 FAs and the possible use of this natural, inexpensive, and safe compound as an alternative treatment for those who are at high risk or have contracted the disease.
The principal source of omega-3 FAs in the human diet comes from eating fresh fish, particularly oily fish, including mackerel, salmon, herring, flounder, cod, and mullet, as shown in Table 2. This happens because most fish foods such as microalgae and other invertebrates are rich in DHA and EPA [45]. Besides this, most microorganisms such as marine protists and dinoflagellates, such as species of Thraustochytrium, Schizochytrium, and Crypthecodinium are rich sources of DHA. On the other hand, microalgae like Phaeodactylum and Monodus are sources rich in EPA [45]. However, we also can find omega-3 amounts in non-marine foods such as cereals, seeds, nuts, and some fruits and vegetables [46, 47].
Omega-3 FAs are PUFAs that are abundantly available in nature. Various forms in which fatty acids exist are free fatty acids (FFAs), ethyl esters, triglycerides, and phospholipids [47, 48]. Dietary lipids after ingestion are hydrolyzed in the intestinal lumen. FFAs and monoglycerides are hydrolysis products that are incorporated into micelles that contain bile salt and get absorbed in enterocytes mostly by passive diffusion [49]. Within the enterocytes, FFAs get incorporated in chylomicrons and via lymphatic circulation will enter the circulation from where lipids are delivered to various organs for further oxidation, storage, or metabolism [49]. Factors like intestinal pH, bile secretion, microorganisms, type of chemical bond, concomitant food intake and presence of some other components like calcium affects the absorption and thus the bioavailability. A diet that consists of FFAs has a higher bioavailability than an ester [50]. Absorption of EPA in triglyceride form is 90% whereas it is 60% in the form of ethyl ester [46]. ALA has low bioavailability due to a higher rate of oxidation, whereas DHA is more bioavailable as a result of being a poor β-oxidation substrate [50]. Binding of long-chain omega-3 FAs in sn-⅓ position to glycerol increases bioavailability by facilitating the bond’s lipase hydrolysis. A study on hamsters conducted by Cholewski and colleagues, showed that DHA when present in sn-2 position facilitates the absorption of fatty acids in the intestine and its tissue incorporation [51]. Metabolism occurs mainly in the liver [52]. ALA gets converted to EPA and DHA by process of desaturation and elongation [53, 54]. Studies show that in healthy young men, approximately 8% of dietary ALA is converted to EPA and 0 – 4% is converted to DHA. Whereas in healthy young females, 21% dietary ALA is converted to EPA and 9% to DHA [55]. ALA with the help of Δ6-desaturase forms Stearidonic acid which then forms Eicosatetraenoic acid via elongation. EPA and DHA are formed by desaturation with the help of Δ5-desaturase [50]. The half-life of EPA is 37 hours and that of DHA is 46 hours [52].
EPA binds and activates PPARα whereas DHA binds and activates PPARγ [56]. When omega-3 FAs are incorporated into the cell membrane, it induces production of eicosanoids and resolvins which then compete with arachidonic acid for the activity of phospholipase A2. FAs are then liberated inside the cytosol and via cyclooxygenase-2 (COX-2), 5-lipoxygenase and thromboxane synthetase activity gets degraded into PGs, LTs, and TX respectively. Thus, omega-3 FA is responsible for decreasing the synthesis of eicosanoids derived from arachidonic acid (AA) like PGI2, PGF2α, PGE2, and LTB4 which are responsible for pro-inflammatory responses and increase the synthesis of LT5 and PGE3 from EPA which have weak effects on inflammation [57, 58].
The guidelines have remained unchanged since 2003 when the American Heart Association recommended consuming at least 0.5 g/day of omega-3 FA to prevent the risk of cardiovascular disease [59]. Despite this, a study conducted by Richter and colleagues showed that more than 90% of American people consumed less than the recommended optimal value for omega-3 FAs in the diet (0.17 g/day) [46]. Aside from maintenance of normal health, omega-3 FAs are also recommended in specific dosages for a variety of inflammatory conditions, as mentioned in Table 3.
Omega-3 FAs belong to a category of supplements known as GRAS (generally recognized as safe). Omega-3 FAs will rarely cause a few mild, non-distressing adverse effects such as unpleasant taste, bad breath, and change in body odor. Some mild gastrointestinal symptoms such as nausea, vomiting, loose stools, and increased stool frequency may be associated with omega-3 FAs. Few patients might report neurologic symptoms such as dizziness and insomnia; however, most symptoms are very mild and self-limiting [50, 57]. Elderly people are at risk of bleeding when they combine long chain PUFA such as fish oil with other anticoagulants such as warfarin and aspirin. The anti-atherosclerotic and anti-lipidemic effects of omega-3 FAs are well known; however, in patients who combine warfarin and fish oil supplements, the risk of bleeding is increased due to inhibition of platelet aggregation. This is the reason which leads to irreversible coagulopathy in elderly patients taking the combination even after suffering blunt head trauma [8, 60].
Omega-3 Fatty Acids and Inflammation
A study conducted by Gutierrez and colleagues showed that omega-3 FA is incorporated throughout the body into the bi-phospholipid layer of the cell membrane of neutrophils and produces different mediators such as prostaglandins, leukotrienes, and maresins. So, if the injury occurs the byproducts of those cell membranes may produce less inflammatory provoking mediators compared to omega-6 FAs, which are more prevalent in the Western diet [58]. Sheppard and colleagues showed that children and adults in the US did not consume sufficient EPA and DHA in their diets. In other terms, Western diets predispose to have a high omega-6/omega-3 ratio which may promote the pathogenesis of many diseases [61, 62]. Further, there is evidence that omega-6 may inhibit the anti-inflammatory effect of omega-3 FAs [63].
Previous studies have been done to clarify the role of omega-3 on anti-inflammatory mechanisms. For example, Saifullah and colleagues carried out a study to know the outcomes of adding EPA and DHA to the diet of hemodialysis patients for 3 months, showing a modest reduction in serum C-reactive protein, which is an inflammatory mediator produced in the liver in response to inflammation [10].
Microorganisms like bacteria, viruses, fungi, protozoa, and worms can all cause infections. Sepsis may occur following infections which can disturb the homeostasis of the body leading to uncontrolled inflammation and ultimately organ failure, shock, and death. A study was done with omega-3 FA lipid emulsion in the cases of sterile peritonitis and murine polymicrobial sepsis demonstrated anti-inflammatory properties of omega-3 FAs. In this study, omega-3 FAs reduced neutrophil infiltration, pro-inflammatory mediators, and classical monocytes while it enhanced non-classical monocyte/macrophage recruitment and efferocytosis in sepsis [64].
Omega-3 FAs include EPA and DHA. Saedisomeolia and colleagues conducted a study to determine the anti-inflammatory properties of DHA and EPA in airway epithelial cells infected with Rhinovirus. In this study, the researchers incubated airway epithelial cells with EPA, DHA, and AA for 24 hours and then infected them with rhinovirus for 48h. They measured IL-6, IL-8, and interferon-gamma-induced protein-10 (IP-10) released by cells using enzyme-linked immunosorbent assay. The investigators found that DHA significantly reduced the release of IL-6 and IP-10 from the cells infected with different strains of rhinovirus. This could be explained by the efficiency of omega-3 FA reducing inflammation by inhibiting AA metabolism to eicosanoids and finally reducing pro-inflammatory cytokines and immune cell function [65].
Omega-3 Fatty Acids and Oxidative Stress
Omega-3 FAs have been found to exhibit antioxidant activity through various mechanisms including upregulating nuclear factor erythroid 2-related factor 2 (NRF2) mediated antioxidant effects, reducing F2 isoprostanes formed during the oxidation of arachidonic acid, inducing PPARγ and modulating toll-like receptor 4 (TLR4) receptors which all lead to a reduction in κβ phosphorylation and thus reduce NF-κβ which in turn reduces inflammatory markers like IL-6, TNFα, and tissue growth factor beta (TGFβ). They also induce mitogen activated protein kinase (MAPK) phosphatases and upregulate glutathione also known as GSH, which is an antioxidant molecule, and upregulate genes responsible for the production of heme-oxygenase, which is cytoprotective. N-3 fatty acids also inhibit lipid peroxidation. They are ultimately metabolized to anti-inflammatory molecules like resolvins, protectins, and maresins [66, 67, 68].
Omega-3 Fatty Acids and Immune System
There is a plethora of ongoing research on the effects of omega-3 FAs and its modifications to the immune system. Omega-3 FAs are considered to be a polyunsaturated fatty acid, which upregulates the activation of immune cells specifically in macrophages, neutrophils, T-cells, B-cells, dendritic cells, natural killer cells, mast cells, basophils, and eosinophils. Omega-3 FA plays a role in the host cellular membrane which regulates membrane fluidity and intricate lipid raft assembling [9]. This is demonstrated in the Figure 2.
Figure 2. How omega-3 fatty acids impact the cellular immune response.Nuclear factor kappa B (NF-κB) is a pro-inflammatory cytokine triggering an inflammatory response via activation of transcription of genes for further pro-inflammatory proteins. Omega-3 fatty acids potentially exert their anti-inflammatory effect via toll-like receptor 4 (TLR4) pathway and G-protein coupled receptor 120 (GPR120) pathway to inhibit the NF-κB and consequently the inflammatory cascade. TAB, TGF-beta activated kinase; TAK, tat-associated kinase; IRAK, interleukin 1 receptor-associated kinase; TRAF6, tumor necrosis factor receptor associated factor 6; TRIAP, TP53 regulated inhibitor of apoptosis; MD2, myeloid differentiation factor 2.
Omega-3 FAs also increase the function of neutrophils which are the first responders of infection in the body. The study showed that omega-3 FAs incorporate phospholipids of the cell membrane of neutrophils and produce different mediators such as prostaglandins, leukotrienes, and maresins. In addition, neutrophils strengthen the immune function by promoting neutrophil migration, phagocytic capacity, and production of reactive free radicals to kill microbes. Omega-3 FAs help activate the function of T cells by promoting antigen-presenting cells (APC), for example, macrophages or dendritic cells. Subsequently, that promotes activation of different subgroups of T cells such as CD4 cells, Th17 cells, and regulatory T cells. B cells are also activated by omega-3 FAs, producing more antibodies utilizing heavy chain immunoglobulin rearrangements and further differentiation. The study claims that omega-3 FAs also increase the population of B cells in the study mice [9]. Omega-3 improves the function of the macrophages by secreting cytokines and chemokines, promoting the ability of phagocytosis, and activating macrophages by polarization [69].
Moreover, the omega-3 FA has appeared to block the activity of NF-κB through decreasing the degradation of the inhibitory subunit of the NF-κB called IkB, in cultured pancreatic cells and human monocyteds. Since NF-κB is responsible for cytokine production in immune cells, by blocking that pathway, its decreasing cytokine storm, and complication [70]. Omega-3 FAs are also known to down-regulate NF-κB. NF-κB is considered to be a transcription factor involved in cell signaling to initiate an inflammatory response by the innate immune system. Furthermore, omega-3 FAs intake upregulates vagal response which in turn down-regulates inflammation and cytokine production. To our understanding omega-3 FAs have multiple effects on the inflammatory response; however, analytic data has not yet recognized its role in critically ill patients. Future research may indicate that supplementation of omega-3 FA fish oils may play a crucial role in SARS-CoV-2 treatment [71].
Omega-3 Fatty Acids and Viral Infection
Omega-3 FAs could be a potential antimicrobial drug with little potential for drug resistance [72]. The metabolites of both omega-3 and omega-6 play an important role in the synthesis of different mediators such as prostaglandins, leukotrienes, thromboxanes, protectins, and resolvins [9]. The study shows that fish oil enhances antiviral response by inducing interferon (IFN) which inhibits viral replication [69]. The anti-inflammatory effect by omega-3 FAs is stronger in DHA compared to that of EPA, and their secretion of cytokines IL-10 is further increased by omega-3. CD8 T cells are responsible for fighting against viruses by inducing the production of different cytokines in the body, such as Tumor Necrosis Factor-alpha (TNF-alpha) and granzyme B. However, the surge of cytokines by CD8 T cells to defend influenza viruses impose unintended lung damage and further deteriorate the clinical outcome. Omega-3 FAs weaken the antiviral response by CD8 T cells and could potentially be used to modulate cytokine responses as antiviral responses, and this process is further explained in Figure 3 [73].
Figure 3. Omega-3 fatty acid acting on different elements of the immune response.Omega-3 fatty acids, through their anti-inflammatory mechanism, inhibit the production of pro-inflammatory mediators like interleukin (IL)-1β, IL-6, tumor necrosis factor (TNF)-α and prevent cytokine storm. Some studies also suggest that they dampen the inflammatory response through regulatory T cells (Treg) differentiation. They also exert an anti-viral effect by enhancing the phagocytic activity of cells of the innate immune system- Neutrophils and Macrophages.
Other Benefits of Omega-3 Fatty Acids
Omega-3 PUFAs include α-linolenic acid (ALA; 18:3 ω-3), stearidonic acid (SDA; 18:4 ω-3), eicosapentaenoic acid (EPA; 20:5 ω-3), docosapentaenoic acid (DPA; 22:5 ω-3), and docosahexaenoic acid (DHA; 22:6 ω-3). The health benefits of omega-3 FA are encompassing and cover a wide range of different organ systems and targets including cardiovascular disease, diabetes, cancer, Alzheimer’s disease, dementia, depression, visual and neurological development, and maternal and child health. Numerous studies have been done that enumerate the reduction in clinical disease rates and mortality rates in patients suffering from a vast array of diseases. Research has been conducted involving humans and omega-3 FA for well over 40 years [74]. Albert and colleagues demonstrated that diets rich in fish and seafood containing high levels of omega-3 PUFAs lead to a statistically significant reduction in mortality from cardiovascular events. The study population included Inuit people who had a diet naturally predominated by seafood and marine life. Men who consumed fish at least once every seven days had a multivariate relative risk of sudden death of 0.48 (95% confidence interval, 0.24 – 0.96; P = 0.04) compared to men who consumed fish less frequently than once per month, demonstrating that diets rich in omega-3’s can reduce the risk of death for men from the cardiovascular event by half [11]. Omega-3 supplementation has also been studied in the setting of ARDS. In a study done in 2015, it was determined that natural antioxidants like omega-3 only lead to a statistically significant decrease in mortality in those suffering from ARDS [75]. Due to the survival rate of COVID-19 patients suffering from ARDS standing at 25%, it is therefore imperative that methods that could potentially provide therapeutic relief or treatment are investigated thoroughly [12]. Although the role of ω-3 supplementation in ARDS needs to be further elucidated, its vital role in reducing reactive oxygen species and pro-inflammatory cytokines, such as TNF-α, IL-1β, IL-6, and IL-8, is widely documented. Therefore, ω-3 PUFAs could be considered for potential interventions for COVID-19 [75].
Omega-3 index, which is the EPA + DHA, is a a percent of total red blood cell fatty acid and is regarded as a new risk factor for death from coronary heart disease (CHD) (Fig. 2). The value of about 8% is estimated to be cardioprotective while that of less than 4% is associated with an increased risk of CHD death [76]. As per our discussion, omega-3 FAs are less prone to the production of different inflammatory markers compared to omega-6 that could initiate a cascade of the cytokine storm. It is evident that the regions with a low index like the USA, few European countries experiencing higher COVID-19 related deaths than those with high index. Omega-3 FAs may play a role in COVID-19 by reducing inflammatory markers and may lower the coronary complications. Figure 4 shows the Global View of Omega-3 Index Levels.
Figure 4. The global view of omega-3 index levels is shown in this figure.The omega-3 index risk zones are as follows: ● High Risk ≤ 4%. ● Intermediate risk = 4 – 8%. ● Low risk > 8%.
Conclusion
There are various benefits of omega-3 FA and taking it as a supplement might be associated with the prevention of the viral entry by changing the composition of fats in the bilipid membrane of cells. Omega-3 FAs, such as DHA and EPA, perform their role by being incorporated in the cell membrane and affecting the clumping of toll-like receptors and thus preventing signals that activate NF-κB and help to ameliorate complications of COVID-19 by producing fewer pro-inflammatory mediators. DHA and EPA are precursors of particles called resolvins D and E, which reduce the proinflammatory mediators thereby reducing pulmonary neutrophils recruitment, increasing apoptosis by macrophages, and subsequently decreasing broncho-alveolar IL-6 production and as a result, decreasing inflammation of the lung. Omega-3 FA plays a role in increasing the phagocytic capacity of macrophages due to the changes in the composition of the cell membrane bilipid layer. Omega-3 FAs also play a role in mediating inflammatory processes and immunomodulation for both innate and acquired immune systems.
It is necessary to understand that the link between omega-3 FA-rich diet and the clinical outcome could be far more complex than previously considered when treating COVID-19 patients. At the very least, this information could serve as the impetus that initiates further conversation and investigation into dietary supplementation that can be considered in both hospitalized patients and patients at home. Several clinical trials are being conducted to investigate the most appropriate treatment for SARS-CoV-2. Due to its anti-inflammatory, immunomodulatory, and other various beneficial properties, omega-3 FA is a natural, inexpensive, and could play a role as a healthier choice of supplement during this ongoing pandemic situation. Future larger randomized blinded clinical trials are warranted and sufficient confirmatory results are required which could further shed light on this topic.
Research shows people with high omega-3 index less likely to die from COVID-19
Pilot study shows positive outcomes for those suffering from COVID-19
Researchers with the Fatty Acid Research Institute (FARI) and collaborators at Cedars-Sinai Medical Center in Los Angeles and in Orange County, CA, have published the first direct evidence that higher omega-3 blood levels may reduce risk for death from COVID-19 infection. The report was published in the journal Prostaglandins, Leukotrienes and Essential Fatty Acids on January 20, 2021.
There are several papers in the medical literature hypothesizing that omega-3 fatty acids should have beneficial effects in patients with COVID-19 infection, but up until now, there have been no published peer-reviewed studies supporting that hypothesis.
This study included 100 patients admitted to the hospital with COVID-19 for whom admission blood samples had been stored. Clinical outcomes for these patients were obtained and blood was analyzed for the Omega-3 Index (O3I, red blood cell membrane EPA+DHA levels) at OmegaQuant Analytics (Sioux Falls, SD). Fourteen of the patients died.
The 100 patients were grouped into four quartiles according to their O3I, with 25% of the patients in each quartile. There was one death in the top quartile (i.e., 1 death out of 25 patients with O3I>5.7%), with 13 deaths in the remaining patients (i.e., 13 deaths out of 75 patients with O3I<5.7%).
In age-and-sex adjusted regression analyses, those in the highest quartile (O3I >5.7%) were 75% less likely to die compared with those in the lower three quartiles (p=0.07). Stated another way, the relative risk for death was about four times higher in those with a lower O3I (<5.7%) compared to those with higher levels.
“While not meeting standard statistical significance thresholds, this pilot study – along with multiple lines of evidence regarding the anti-inflammatory effects of EPA and DHA – strongly suggests that these nutritionally available marine fatty acids may help reduce risk for adverse outcomes in COVID-19 patients. Larger studies are clearly needed to confirm these preliminary findings,” said Arash Asher, MD, the lead author on this study.
Agreeing with Dr. Asher, cardiology researcher and co-developer with Dr. Harris of the Omega-3 Index, Clemens von Schacky, MD, (CEO, Omegametrix GmbH, Martinsried, Germany, and not involved with the study) said, “Asher et al have demonstrated that a low Omega-3 Index might be a powerful predictor for death from COVID-19. Although encouraging, their findings clearly need to be replicated.”
Omega-3 expert James H. O’Keefe, Jr., MD, (Director of Preventive Cardiology, Saint Luke’s Mid America Heart Institute, Kansas City, MO, and also not involved with the study) observed, “An excessive inflammatory response, referred to as a ‘cytokine storm,’ is a fundamental mediator of severe COVID-19 illness. Omega-3 fatty acids (DHA and EPA) have potent anti-inflammatory activities, and this pilot study provides suggestive evidence that these fatty acids may dampen COVID-19’s cytokine storm.”
The FARI research team is currently seeking funding to expand upon these preliminary observations. Individuals and organizations that want to support this research are encouraged to visit FARI’s donations page.
The Fatty Acid Research Institute (FARI) is a non-profit research and education foundation. FARI was founded in order to accelerate discovery of the health effects of fatty acids, most notably, the long chain omega-3 fatty acids EPA and DHA. FARI researchers and scientists will focus single-mindedly on publishing high-quality research studies on the multiple relationships between fatty acid levels and human (and animal) health outcomes. These studies will improve the ability to predict risk for disease, and more importantly, suggest ways to reduce risk by changing our diets and/or supplementation regimens.
ChooseLife Notes:
Again, this builds the picture out, that likelihood of acquiring and gradient of infection are both clearly linked to multiple biological processes, these processes are demonstrably underpinned in many instances, or enhanced greatly in others, by the intelligent adaptation of diet to include plentiful amounts of Omega3, my preference is FlaxOil via Budwig, it adds so many extra facets, partcularly in Mucin production and intestinal fauna well-being (but I am perhaps biased a little as a vegetarian!).
Understanding how dietary components alter the healthy baseline colonic microenvironment is important in determining their roles in influencing gut health and gut-associated diseases. Dietary flaxseed (FS) has demonstrated anti-colon cancer effects in numerous rodent models, however, exacerbated acute colonic mucosal injury and inflammation in a colitis model. This study investigates whether FS alters critical aspects of gut health in healthy unchallenged mice, which may help explain some of the divergent effects observed following different gut-associated disease challenges. Four-week-old C57Bl/6 male mice were fed an AIN-93G basal diet (BD) or an isocaloric BD+10% ground FS diet for 3 weeks. FS enhanced colon goblet cell density, mucus production, MUC2 mRNA expression, and cecal short chain fatty acid levels, indicative of beneficial intestinal barrier integrity responses. Additionally, FS enhanced colonic regenerating islet-derived protein 3 gamma (RegIIIγ) and reduced MUC1 and resistin-like molecule beta (RELMβ) mRNA expression which may indicate altered responses in regulating microbial defense and injury repair responses. FS diet altered the fecal microbial community structure (16S rRNA gene profiling), including a 20-fold increase in Prevotella spp. and a 30-fold reduction in Akkermansia muciniphila abundance. A 10-fold reduction in A. muciniphila abundance by FS was also demonstrated in the colon tissue-associated microbiota (quantitative PCR). Furthermore, fecal branched chain fatty acids were increased by FS, indicative of increased microbial-derived putrefactive compounds. In conclusion, consumption of a FS-supplemented diet alters the baseline colonic microenvironment of healthy mice which may modify subsequent mucosal microbial defense and injury-repair responses leading to altered susceptibility to different gut-associated diseases.
Flax seed oil and flax seed meal are good sources of omega-3 fatty acids. The objective of this study was to explicate the effects of feeding flax seed oil and flax seed meal on AOM-induced aberrant crypt foci (ACF) in Fisher 344 male rats. Following an acclimatization period, rats were divided into six groups and fed AIN 93G diet Control (C), C+7 and 14% soybean oil (SBO), C+7 and 14% flax seed oil (FSO) and C+10 and 20% flax seed meal (FSM). All rats received 16 mg/kg body weight of AOM at 7 and 8 weeks of age. The rats were euthanized with CO2 at 17 weeks of age. FSM and FSO reduced the incidence of ACF which are putative precursor lesions in the development of colon cancer in the distal colon by 88% and 77%, in the proximal colon by 86% and 87% with a total reduction of 87.5% and 84%, respectively. Glutathione-S-transferase (GST) activities were significantly (P<0.05) higher in rats fed C+7 and 14% FSO and C+10 and 20% FSM, as compared to rats fed C+SBO diets. Results of this study showed that FSO and FSM reduced the incidence of AOM-induced ACF formation and may therefore be effective chemopreventive agents.
The chronic and low-grade inflammation induced by obesity seem to be the “first hit” to retinopathy associated to diabetes type 2. Herein, we hypothesized that omega-3 fatty acids from flaxseed oil enriched diet disrupt the pro-inflammatory status in the retina, protecting against retinopathy development. For eight weeks under a high-fat diet (HF), several physiological parameters were monitored to follow the metabolic homeostasis disruption. After this period, mice were treated with a HF substituted in part of lard by flaxseed oil (FS) for another eight weeks. Food behavior, weight gain, glucose and insulin sensitivity, electroretinography, RT-qPCR and western blots were carried out. The HF was able to induce a pro-inflammatory background in the retina, changing IL1β and TNFα. VEGF, a master piece of retinopathy, had early onset increased also induced by HF. The FS-diet was able to decrease inflammation and retinopathy and improved retinal electro stimuli compared to HF group. GPR120 and GPR40 (G Protein-Coupled Receptors 120 and 40), an omega-3 fatty acid receptors, were detected in the retina for the first time. FS-diet modulated the gene expression and protein content of these receptors. Thus, unsaturated fatty acids protect the retina from diabetes type 2 mice model from disease progression.
The aim of this article is to examine the effect of omega-3 (ω-3) long-chain polyunsaturated fatty acids (LCPUFAs) intake on retinopathy of prematurity (ROP) by reviewing the experimental and clinical trials conducted on animal models and infants. LCPUFAs demonstrate cytoprotective and cytotherapeutic actions contributing to a number of anti-angiogenic and neuroprotective mechanisms within the retina. Their intake appears to have a beneficial effect on ischemia, oxidative stress, inflammation and cellular signaling mechanisms, influencing retinal cell gene expression and cellular differentiation. ω-3 LCPUFAs may modulate metabolic processes that activate molecules implicated in the pathogenesis of vasoproliferative and neurodegenerative retinal diseases such as ROP.
ChooseLife : Arya Fosh, my wonderful daughter, was born at 24 weeks and 0 days. She was born with a host of arrival injuries, she developed Retinopathy Of Prematurity due to the Oxygen support required through her Chronic Lung Disease (as our daughter was prone to desaturation often the Oxygen would be left high to stabilise and leave her on 100%, which our first Consultant at Portsmouth NICU had previously said (to a Junior Doctor in my earshot) was criminal and had stuck in my mind, due to the damage it does to the eyes development of the delicate blood vessels linking the retina, Stevie Wonders ‘blindness’ came from this dynamic), so I became very prickly with nurses more interested in chatting than ensuring that she did not sit at 100% on higher supplemental Oxygen, once leading to a full argument with a lazy nurse chatting about TV whilst my daughters eyes were being harmed by her shameful actions, she had the gall to say our concerns were unfounded as she was too old to develop ROP, she hadn’t even read the notes which stated she was Grade 2 progressive to grade 3! This was at Dorchester SCBU).
During this time her mother was plied with a plentiful supply of Omega3 via Fermented Cod Liver Oil and Butter Oil (Royal Ice), based on the studies of the Weston A. Price foundation. The 2:1 ratio of Omega6/Omega3 plus the Vitamin A content (in a 5:1 ratio with Vitamin A to D). This nourished our daughter via her mothers expressing milk, I am in no doubt.
We were told our daughter needed to be transferred to Southampton NICU, as her ROP was measured as grade 2 progressing towards grade 3 and eye surgery was required, this was another traumatic experience, as transferring our daughter caused desaturations and posed risk to her stability and life. Once we arrived, we were met by the Surgeon who performed a pre-surgery exam, the Surgeon reported to us that contrary to previous analysis our daughter was in fact healing and would not require surgery, a huge huge relief for us.
The surgeon stated that we must have good genes for Arya to be regressive and her eye health looked good, inside I knew it was the dietary support we had engaged which was at play, this study only goes to confirm this belief.
We were told she would never be 20:20 due to this dynamic, however she was classed as 20:20 by the age of 3!
Praise be to Omega 3! Arya will never be without it (though now via ultra fresh Flaxoil blended with Quark 1:2, as per Johanna Budwig’s sensational research).
The current coronavirus disease 2019 (COVID-19) pandemic presents a global challenge for managing acutely ill patients and complications from viral infection. Systemic inflammation accompanied by a “cytokine storm,” hemostasis alterations and severe vasculitis have all been reported to occur with COVID-19, and emerging evidence suggests that dysregulation of lipid transport may contribute to some of these complications. Here, we aim to summarize the current understanding of the potential mechanisms related to COVID-19 dyslipidemia and propose possible adjunctive type therapeutic approaches that modulate lipids and lipoproteins. Specifically, we hypothesize that changes in the quantity and composition of high-density lipoprotein (HDL) that occurs with COVID-19 can significantly decrease the anti-inflammatory and anti-oxidative functions of HDL and could contribute to pulmonary inflammation. Furthermore, we propose that lipoproteins with oxidized phospholipids and fatty acids could lead to virus-associated organ damage via overactivation of innate immune scavenger receptors. Restoring lipoprotein function with ApoA-I raising agents or blocking relevant scavenger receptors with neutralizing antibodies could, therefore, be of value in the treatment of COVID-19. Finally, we discuss the role of omega-3 fatty acids transported by lipoproteins in generating specialized proresolving mediators and how together with anti-inflammatory drugs, they could decrease inflammation and thrombotic complications associated with COVID-19.
Department of Clinical and Experimental Medicine, University of Foggia, Italy
2. Department of Experimental Medicine, Section of Human Physiology and Unit of Dietetics and Sports Medicine, Università degli Studi della Campania Naples, Italy
3. Department of History, Society and Studies on Humanity, University of Salento, Lecce Italy
4. Department of Law, Forensic Medicine, Magna Graecia University of Catanzaro, Italy
5. Department of General Surgery and Medical-Surgical Specialties, University of Catania, Italy
6. Department of Mental Health, Physical and Preventive Medicine, Clinic of Child and Adolescent Neuropsychiatry, Università degli Studi della Campania Italy
7. Department of Education Sciences, Psychology and Communication, University of Bari, Italy
Abstract
Background: On the 31 December 2019, the World Health Organization (WHO) was informed of a cluster of cases of pneumonia of unknown origin detected in Wuhan City, Hubei Province, China.
The infection spread first in China and then in the rest of the world, and on the 11th of March, the WHO declared that COVID-19 was a pandemic. Taking into consideration the mortality rate of COVID-19, about 5–7%, and the percentage of positive patients admitted to intensive care units being 9–11%, it should be mandatory to consider and take all necessary measures to contain the COVID-19 infection.
Moreover, given the recent evidence in different hospitals suggesting IL-6 and TNF-α inhibitor drugs as a possible therapy for COVID-19, we aimed to highlight that a dietary intervention could be useful to prevent the infection and/or to ameliorate the outcomes during therapy. Considering that the COVID-19 infection can generate a mild or highly acute respiratory syndrome with a consequent release of pro-inflammatory cytokines, including IL-6 and TNF-α, a dietary regimen modification in order to improve the levels of adiponectin could be very useful both to prevent the infection and to take care of patients, improving their outcomes.
1. Background
On the 31 December 2019, the World Health Organization (WHO) was informed of a cluster of cases of pneumonia of unknown origin detected in Wuhan City, Hubei Province, China. About one month later (on 8 January 2020), the Chinese authorities declared the identification of a new type of coronavirus, informing the WHO a few days later that the outbreak was associated with exposure in a seafood market in Wuhan City.
The infection spread firstly in China and then in the rest of the world, and on the 11th of March, the WHO declared that COVID-19 was a pandemic.Coronaviruses (CoVs) belong to the subfamily Orthocoronavirinae in the family of Coronaviridae in the order Nidovirales, and this subfamily includes α-coronavirus, β-coronavirus, γ-coronavirus, and delta-coronavirus [1].
Coronaviruses primarily cause enzootic infections in birds and mammals and, in the last few decades, have shown to be capable of infecting humans as well [2]. In human infections with highly virulent respiratory viruses—such as avian influenza H5N1, H7N9, Severe Acute Respiratory Syndrome (SARS) coronavirus, and Coronavirus Disease-19 (COVID-19)—immunopathogenesis caused by the overproduction of pro-inflammatory cytokines may play an essential role in disease progression and mortality [3].
Several recent studies have reported that COVID-19 caused the destruction of the pulmonary parenchyma, including interstitial inflammation and extensive consolidation, similarly to the previously reported coronavirus infection [4,5]. During coronavirus infection, it was observed that the lungs increased in weight, with a mild pleural effusion of clear serous fluid, named pulmonary edema, and extensive consolidation [6,7]. In some areas, there was interstitial thickening, with mild-to-moderate fibrosis, but a disproportionately sparse infiltrate of inflammatory cells (mainly histiocytes, including multinucleated forms, and lymphocytes) [8]. A dilatation of the airspaces was observed, as was focal honeycombing fibrosis. An intra-alveolar organization of exudates was described, and the formation of granulation tissues in the small airways and airspaces was reported. These lesions were typically located in the sub-pleural region, and the cellular component mainly consisted of histiocytes, as reported in a previous paper [9]. Xu et al. described in their case report the pathological findings of COVID-19 associated with acute respiratory distress syndrome. At the X-ray investigation, they detected a rapid progression of bilateral pneumonia.
The biopsy samples were taken from the lung; the histological examination showed bilateral diffuse alveolar damage with cellular fibromyxoid exudates [6].Considering that the mortality rate of COVID-19, about 5–7% [10], and the percentage of positive patients admitted to intensive care units being 9–11% [11], it should be mandatory to consider and take all necessary measures intended to contain the viral infection.
A recent study analyzed the data of 150 COVID-19 patients, with the aim of defining the clinical predictors of mortality. The results obtained from this study suggest that COVID-19 mortality might be due to virus-activated “cytokine storm syndrome”, considering that the plasma levels of IL-6 were higher in deceased patients compared to in discharged subjects [12].Considering that a detailed study has not been performed on the immunological response to COVID-19, the only way to discuss this thematic is to refer to previous knowledge about SARS-CoV and MERS-CoV. The first response is obtained through pattern recognition receptors (PRRs) including C-type lectin-like receptors, Toll-like receptors (TLR), NOD-like receptors (NLR), and RIG-I-like receptors (RLR). Moreover, several inflammatory factors are expressed such as IL-6 and TNF-α; moreover, the synthesis of type I interferons (IFNs) is activated, and these exert their actions against virus diffusion, accelerating macrophage phagocytosis [13] (Figure 1).
Figure 1. The main immunological response to COVID-19.
In the light of these considerations and the recent evidence in different hospitals suggesting IL-6 and TNF-α inhibitor drugs as a possible therapy for COVID-19, this review aims to highlight how a dietary intervention could be useful to prevent the infection and/or to ameliorate the outcome during therapy.
2. The Pivotal Role of IL-6 and TNF-α in Lung Infections
The first laboratory report about COVID-19 patients indicated several parameters that were found to be altered in blood samples; for example, D-dimer, neutrophil count, blood urea, and creatinine levels were significantly higher. In the same way, several cytokines such as IL-6 and TNF-α were overexpressed, indicating the immune status of the patients [14].IL-6 represents pro-inflammatory signaling produced by adipose tissue; for this reason, this endocrine cytokine could be important in regulating the host response during acute infection [15].
Several papers have described the essential role of IL-6 in generating a proper immune response during different kinds of viral infection in the pulmonary tract. Others link this cytokine to an exacerbation of viral disease. These latter findings support the hypothesis that IL-6 upregulation during viral infections may promote virus survival and the exacerbation of the clinical disease [16,17].
Indeed, IL-6 has a pleiotropic function, and it is produced in response to tissue damage and infection. In particular, at the pulmonary level, innate and adaptative immune cell proliferation is strongly influenced by this cytokine. After targeting its specific receptor, IL-6 starts a cascade of signaling events mainly associated with the JAK/STAT3 activation pathway, promoting the transcription of multiple downstream genes related to cellular signaling processes, including cytokines, receptors, adaptor proteins, and protein kinase [15].
Furthermore, it has been reported that IL-6 is an essential factor for the survival of mice with a viral infection. This cytokine promotes the optimal regulation of the T-cell response, inflammatory resolution, tissue remodeling promoting lung repair, cell migration, and the phagocytic activities of macrophages, as well as preventing virus-induced apoptosis in lung epithelial cells.
However, experimental scientific evidence also suggests potential adverse consequences that increased levels of IL-6 might have on the cellular immune response against viruses. In this context, different possible mechanisms involving this cytokine might affect viral clearance, ultimately favoring the establishment of a persistent viral state in infected hosts [18,19].
Tumor necrosis factor is a cell-signaling protein (cytokine) involved in systemic inflammation, released predominately from macrophages, but it is also released from a variety of other immune cells. It has been well described that during infection with the influenza virus, the expression of TNF-α in lung epithelial cells was higher, exerting powerful anti-influenza virus activity [20].
In an animal model, it has been demonstrated that TNF-α plays a pivotal role in the development of pulmonary fibrosis. TNF-α signals via two receptors, TNF-RI and TNF-RII; the first receptor (TNF-RI) promotes intracellular signaling involving c-Jun N-terminal kinase (JNK) and nuclear factor (NF)-κB, while the other receptor, TNF-RII, promotes TNF-RI–dependent cell death, without directly inducing apoptosis. Although both receptors are broadly expressed, it is known that the majority of inflammatory signaling is elicited through TNF-RI [21].
In an in vitro model, it has been described that serine/threonine kinases can phosphorylate TNF-RI and its molecules, preventing tyrosine phosphorylation [22,23,24].In patients with COVID-19, the high serum levels of IL-6 and TNF-α are negatively correlated to T cells; contrariwise, it has been demonstrated that T cell levels were restored by reducing IL-6 and TNF-α concentrations [25]. These findings suggested that these cytokines could represent important targets of anti-COVID-19 therapies.
3. Adiponectin Function in Lung Infections
Through the secretion of adipokines, adipose tissue participates in the regulation of several pathophysiological processes in many organs and tissues. Among the adipokines, adiponectin is the most relevant. Adiponectin is one of the most abundant circulating adipocytokines, accounting for 0.01% of total serum protein. Adiponectin is an important regulator of cytokine responses, and this effect is isoform-specific. It is involved in a wide variety of physiological processes, including energy metabolism, inflammation, and vascular physiology. These effects are mediated by two atypical, widely expressed seven-transmembrane receptors, AdipoR1 and AdipoR2 [26]. Adiponectin has beneficial effects in cardiovascular systems and blood vessels, protecting these tissues through the inhibition of pro-inflammatory and hypertrophic responses and stimulation of endothelial cell responses [27].
Adiponectin circulates as three different isoforms (low molecular weight—LMW, medium molecular weight—MMW, and high molecular weight—HMW) [28].Infectious diseases are characterized by an increased production of adiponectin. Several papers suggest that adiponectin may be related to disease activity and/or severity in different conditions such as rheumatoid arthritis, osteoarthritis, and systemic lupus erythematosus. Since adiponectin has been found to display both pro- and anti-inflammatory activities, controversial findings have been observed regarding the role of total adiponectin in systemic autoimmune and inflammatory joint diseases. For this reason, the relative contribution of each adiponectin isoform to the inflammatory response and joint and/or tissue damage requires further study [29].
It is reported that adiponectin is regulated by transcription factors in adipose tissue, such as peroxisome proliferator-activated receptor-γ (PPAR-γ) [30]. During viral infections, it has been reported that the role of the predisposition of hosts is also important, as well as their state of health and nutrition. Indeed, it is well known that white adipose tissue is considered an endocrine source of biologically active substances with local and/or systemic action, called adipokines.
The inappropriate secretion of adipokines seems to participate in the pathogenesis of obesity-related diseases, including endothelial dysfunction, inflammation, and atherosclerosis [31,32,33].The biological function of adipokines in lung diseases seems to be mainly related to the inflammatory process. In particular, the intercorrelation between adipose tissue and the lung has become evident as the involvement of adiponectin has been demonstrated in several lung diseases such as Chronic Obstructive Pulmonary Disease (COPD), emphysema, and cancer [34]. In fact, with specific regard to COPD, a low-grade inflammatory state has been demonstrated [35,36,37].
Moreover, increasing evidence suggests that adiponectin also exerts a crucial role in the vascular endothelium, maintaining vascular homeostasis and protecting against vascular dysfunctions. Altogether, these findings support the anti-inflammatory role of adiponectin in COPD and, in general, in other lung diseases [38].The critical role of adiponectin in the pathophysiological conditions of the lung is also supported by the modulation of AdipoRs with the downregulation of AdipoR2. It has been described that the adiponectin oligomerization state is altered in COPD; moreover, the presence of AdipoR1 and AdipoR2, with a lower expression of AdipoR2 compared to AdipoR1, in lung tissue [39] has been demonstrated. The low expression of AdipoR2 could suggest a specific role of this receptor, mainly implicated in adiponectin’s effects on inflammation and oxidative stress. Mainly, it has been observed that higher levels of adiponectin are associated with a significant and specific increase in HMW adiponectin, representing the most biologically active forms. Thus, HMW adiponectin increases IL-6 secretion in human monocytes and human monocytic leukemia cell lines but does not suppress lipopolysaccharide (LPS)-induced IL-6 secretion. Byn contrast, LMW adiponectin reduces LPS-mediated IL-6 release and also stimulates IL-10 secretion [40].
Furthermore, several in vitro studies have demonstrated that adiponectin in the A549 adenocarcinoma human alveolar basal epithelial cell line has an essential apoptotic effect and also reduces the production of pro-inflammatory cytokines such as TNF-α, blocking NF-κB nuclear translocation [41,42].Indeed, adiponectin can reduce innate and adaptive immune cell proliferation and polarization, also blocking the production of pro-inflammatory cytokines such as TNF-α, IL-2, and IL-6, and enhancing that of anti-inflammatory cytokines such as IL-10, with a decrease in the phosphorylation of AMPK, p38, ERK1/2, and c-JNK [43,44,45,46]. Data from in vitro studies on lung cells were consistent with an anti-inflammatory function of adiponectin, and adiponectin-deficient mouse models developed lung function impairments and systemic inflammation [47].
The possible role of adiponectin in inflammatory pulmonary diseases, such as asthma and chronic obstructive pulmonary disease (COPD), and in critical illnesses has been the subject of recent investigations. Particularly, the HMW isoform has a specific role in pulmonary diseases and critical illnesses, even if its role should be better clarified [48,49].
An interesting study reported that systemic adiponectin concentrations in humans fall during the acute phase of lung infection: particularly, during the early phase, the pro-inflammatory state is generated by the high systemic TNF-α and IL-6 concentrations, with the subsequent inhibition of adiponectin production. Contrariwise, it has been described that the reduction in TNF-α and IL-6 factors generates a corresponding bounce-back in systemic adiponectin concentrations [50].
Although it is still unclear whether the modulation of systemic adiponectin or its signaling pathways has any therapeutic benefit in pulmonary or critical illnesses, it may serve as a novel therapeutic or preventative tool for these illnesses in the future. One obvious pharmaceutical treatment would be the exogenous administration of adiponectin by the inhalational or intravenous route. Although this has been tried in mouse models [51], the problems to be overcome prior to human administration include establishing what the biologically active molecule is and what role post-translational modifications have upon its function, and the associated difficulties in generating biologically active molecules on a large scale.
Considering the difficulty linked to the direct administration of adiponectin, in the last few years, other drugs have been used that indirectly improve adiponectin production. For example, a synthetic ligand of peroxisome proliferator-activated receptors can increase adiponectin mRNA in adipocytes, improving the production and secretion of adiponectin [52,53,54,55]. Moreover, other drugs such as fibrates can increase systemic adiponectin levels by enhancing PPAR-γ activity [56,57]. Another way to improve adiponectin levels is the use of angiotensin converting enzyme inhibitors [58,59,60]. Furthermore, it is possible to stimulate adipocyte differentiation [61] and the activation of PPAR [62].
Finally, it has been described that calcium channel blockers [63] and a central-acting anti-hypertensive agent [64] also increase systemic adiponectin concentrations [65]. The possibility to improve the action of adiponectin through diet is intriguing; it has been described that nutritional interventions may help to regulate systemic adiponectin concentrations. In an animal model, it has been demonstrated that a diet with a high concentration of polyunsaturated fatty acids and supplemented with ω-3 can improve the plasma levels of adiponectin, increasing gene expression [66]. On the other hand, in humans, adiponectin levels are positively associated with a healthy lifestyle and the Mediterranean diet, even if the mechanisms of action are not completely known [66]. Finally, in light of these considerations, in COVID-19 therapy, it could be very useful to combine drug therapy with a specific diet regimen.
4. ω-3 PUFAs and Lung Infections
Another important mediator involved in the immune response and influenced by nutrition are fatty acids, in particular, ω-3 PUFAs [67,68]. In fact, during bacterial and viral infections, they are able to act on immune cells and regulate diverse inflammatory processes. ω-3 PUFAs are known to have anti-inflammatory properties and play an essential role in the resolution of inflammation [69].
In several lung infections, the administration of PUFA can ameliorate the outcome of the patient in acute pneumonia. Sharma et al. reported in their study that the dietary supplementation of ω-3 PUFA can exert an overall beneficial effect against acute pneumonia through the upregulation of the host’s specific and nonspecific immune defenses [70]. ω-3 polyunsaturated fatty acids (PUFA, ω-3-fatty acids), the key components of fish and flaxseed oils, are increasingly consumed by the public because of their potential health benefits and can be used clinically for the treatment of metabolic, cardiac, inflammatory, and autoimmune diseases [71].
However, numerous studies have shown that these compounds are immunoregulatory and immunosuppressive and thus may increase susceptibility to infection. While reports suggest that ω-3 PUFAs may have beneficial effects against extracellular pathogens, few studies have been performed on systemic viral infections in mammals. Jones and Roper described in their study that a diet rich in ω-3 PUFAs did not significantly lower survival of the vaccinia virus infection, at least with short-term (~6 week) feeding in mice [71].
ω-3 PUFAs are metabolized into various mediators possessing anti-inflammatory properties such as resolvins and protectins. It is known that ω-3 PUFAs can reduce NF-κB activation by preventing nuclear p65 NF-κB translocation. Furthermore, ω-3 PUFAs minimize the activation of ERK1/2 MAPK, also reducing COX-2 production. The ω-3 PUFA-derived lipid mediator could markedly attenuate influenza virus replication via the RNA export machinery. In addition, the treatment of protectin D1 with peramivir could completely stop mouse mortality [72].
ω-3 supplementation was previously studied in Acute Respiratory Distress Syndrome (ARDS). Singer and Shapiro suggested that the enteral administration of natural antioxidant substances could improve oxygenation and clinical outcomes in ICU patients [73]. A systematic review performed in 2015 reported a positive effect only for patients suffering from ARDS with high mortality [74]. A more recent meta-analysis highlighted the importance of clinical trials in order to clarify the use of ω-3 fatty acids and antioxidants in patients with ARDS to ascertain the positive effects in order to reduce the lengths of ICU stays and the numbers of days spent on ventilators [75].
Although the role of ω-3 supplementation in ARDS should be better clarified, its pivotal role in reducing reactive oxygen species and pro-inflammatory cytokines, such as TNF-α, IL-1β, IL-6, and IL-8 [76], is well known.Therefore, ω-3 PUFAs, including protectin D1, which is a novel antiviral drug, could be considered for potential interventions for COVID-19.
5. Other Dietary Constituents and Lung Infections
As previously described, other dietary constituents can be used to improve the patients’ outcomes during lung infection, regulating the inflammatory response. Among these, antioxidants play an important role in protecting lung cells against viruses and bacteria. Viral infection leads to an increase in the intrapulmonary oxidative burden. In many diseases, the balance between oxidants and antioxidants (redox balance) is altered, with severe consequences [77].
The pathophysiological mechanisms by which free radicals generate various types of stress—such as oxidative, nitrative, carbonyl, inflammatory, and endoplasmic reticulum stress—lead to lung inflammation and an altered lung immune response. In this scenario, dietary antioxidants may play an important role against lung oxidative stress [77].
Several studies reported the protective role of the antioxidants in lung infection and in lung inflammation [78,79].In particular, vitamin C, polyphenols, and flavonoids can play a protective role in lung infections, being immune modulators and inflammatory mediators. Indeed, as reported by Carr et al., during infection, vitamin C levels may become depleted; for this reason, vitamin C supplementation can attenuate infection. Based on this evidence, these authors suggested a clinical trial with vitamin C infusion for the treatment of severe COVID-19 patients [80].
Among polyphenols, epigallo-catechin 3 gallate (EGCG) is the most potent ingredient in green tea and exhibits antibacterial, antiviral, antioxidative, anticancer, and chemo-preventive activities. Recently, numerous studies have investigated the protective effects of EGCG against asthma and other lung diseases such as COPD and lung pneumonia. EGCG may suppress inflammation and inflammatory cell infiltration into the lungs of asthmatic mice, and may also inhibit epithelial-mesenchymal transition EMT via the PI3K/Akt signaling pathway through upregulating the expression of phosphatase and tensin homolog (PTEN), both in vivo and in vitro [81].
Moreover, flavonoids can be used to attenuate lung injury in mice; it has been reported that they inhibit influenza virus and Toll-like receptor signaling, blocking NF-κB translocation [82].Therefore, as summarized in Table 1, supplementation with vitamin C, flavonoids, and polyphenols can be tested in COVID-19 patients, both in order to prevent viral infection and to improve patients’ outcomes.
Table 1. The principal antioxidants involved in lung infection and the immune-inflammatory response.
6. Discussion and Conclusions
During pulmonary infections, and particularly in COVID-19 patients, intracellular signaling leads to the production of pro-inflammatory cytokines, such as TNF-α and IL-6, which act in concert with chemoattractants, such as CXCL1 and CXCL2, to recruit polymorphonuclear leukocytes (PMNs) to the lungs, killing pathogens but generating fibrosis [83].
Another important consideration during COVID-19 infection is related to the modification of the secretory products of the upper and lower airways, which usually include mucin and pulmonary surfactant. During infection, mucin production is upregulated, with the function of preventing microbes from binding to and infecting epithelial cells [84].
The primary source of phospholipids (PLs) in the lung is pulmonary surfactant, synthesized and released by alveolar epithelial type II cells. The surfactant contains approximately 80–90% PLs, with fatty acid chains that can be oxidized during different challenges in the lung [85]. The oxidation of these PLs in the lung can occur in the setting of an increased oxidative stress situation, such as infection and inflammation [86]. The immune effects of oxidized phospholipids oxPLs during infectious diseases are inevitably dictated by the balance among activation, degradation, and scavenging. It has been shown that oxPLs are generated in the lung during several pulmonary infections, including influenza and avian influenza (H5N1), as well as SARS coronavirus, even if the mechanisms of action are not well known [87,88,89].
As reported by Imai et al., oxPL-induced inflammation is mediated by TLR4 and TRIF, driving an increase in IL-6 production [89]. It is intriguing to consider that oxPL-dependent defects in phagocytosis and ROS generation may lead to an increased susceptibility to respiratory infections [90]. Cholesterol is the major neutral lipid in pulmonary surfactant, in which it is thought to promote the spreading, mobility, and adsorption of surfactant films [91].
As previously documented, modulating adiponectin levels can be considered an important way to reduce cytokines levels; in this way, the adverse effects related to the COVID-19 infection should be attenuated. It is well described in animal models that the consumption of hyperlipidemic diets, rich in saturated fat, reduces the levels of adiponectin, while diets rich in polyunsaturated fatty acids and supplemented with ω-3 PUFA increase adiponectin levels, reducing pro-inflammatory cytokines [66].Innate and adaptive immune responses are influenced not only by oxPLs and cholesterol but also by the fatty acid profiles of tissues in response to pharmacological agents and diet [92].
Several studies performed in animal models demonstrated how ω-3 PUFA uptake into the lung tissue influences outcomes associated with infection, promoting the resolution of inflammation [93]. In another study, ω-3 PUFAs reduced the levels of PMNs and lowered IL-6 levels in lung infections [94]. These positive effects remain controversial; for example, Jones and Roper reported that in their experimental model, no statistically significant differences were found among the diet regimens, with and without ω-3 PUFAs, with respect to the susceptibility of mice to viral infection, morbidity, viral organ titers, recovery time, or mortality [71].
In conclusion, it is well known that general treatments are very important to enhance the host immune response against RNA viral infection. In addition, the immune response has often been shown to be weakened by inadequate nutrition in many model systems as well as in human studies. However, the nutritional status of the host, until recently, has not been considered as a contributing factor to the emergence of viral infectious diseases. The recent reports about the pathogenesis of COVID-19 suggested that one of the most important consequences of this infection is the cytokine storm syndrome [95], which could be strictly linked with coagulopathy, generating acute pulmonary embolism caused by in-situ thrombosis [96,97]. Therefore, a great number of clinical trials are ongoing to define a useful therapy to attenuate cytokine storms [98].For these reasons, an adequate ω-3 PUFA intake may be a valid strategy against viral infection.
Indeed, following the recommended intake of ω-3 PUFA, in the range of 0.5% and 2% of total calories (250 mg/day), may be important to protect against an excessive inflammatory response, also reducing IL-6 levels. This theory found important support in a recent study that demonstrated that ω-3 PUFA-derived lipid mediator protectins can suppress influenza virus replication through a mechanism that blocks the export of viral mRNA. Moreover, Imai demonstrated that this mediator can be used in combination with the antiviral peramivir, even at late time points in infection [99].
Nevertheless, the efficacy of ω-3 PUFAs at the clinical level is under investigation; for example, Hecker et al. described a beneficial effect for a diet regimen with ω-3 PUFAs, describing that the pro-inflammatory cytokine levels decreased after this diet regimen [100]. The suggested positive role in the outcome and prevention of the COVID-19 infection is summarized in Figure 2.
Figure 2. Adiponectin and ω-3 PUFAs reduce the lung inflammation that occurs following coronavirus infection, reducing IL-6 production, ERK1/2, and COX-2 activation and the nuclear translocation of NF-κB.
In addition, adiponectin plays a role in lung diseases and obesity; in the development and progression of lung disease and cancer, a pathogenic role of adiponectin was defined by both in vivo and in vitro studies.
Recently, immunometabolic pathomechanisms have been identified as important factors determining and modulating lung function and disease. Particularly, adiponectin levels have been found to be greater in patients with COPD compared with in control patients, and adiponectin-deficient mice are protected from several lung diseases [101].
Moreover, it has been reported that adherence to the Mediterranean diet was associated with an increase in adiponectin levels, improving cardiovascular system functionality [102], particularly in elderly people [103]. These findings are only apparently contradictory to the first data about the mortality rate from COVID-19 infections in the Mediterranean area (such as in Italy and Spain) [104].
First of all, the data have been referred only to the tested population; moreover, it is well described that the presence of several comorbidities such as hypertension, diabetes, and cardiovascular diseases severely influenced the mortality rate reported in this area [105].
All these comorbidities can be counteracted with a correct dietary regimen. Therefore, both adiponectin and ω-3 PUFAs appear to be attractive biomarkers for monitoring lung disease progression.
Finally, considering that the COVID-19 infection can generate a mild or highly acute respiratory syndrome with a consequent release of pro-inflammatory cytokines, including IL-6 and TNF-α, a modification of the dietary regimen in order to improve the levels of adiponectin could be very useful both to prevent the infection and to take care of the patients, improving their outcomes.
Given the similar pathway of action, it can be hypothesized that adiponectin and ω-3-PUFA could be used as real drugs to reduce inflammation, reducing both IL-6 and TNF-α levels as well as ameliorating the lung damage that occurs following coronavirus infection.