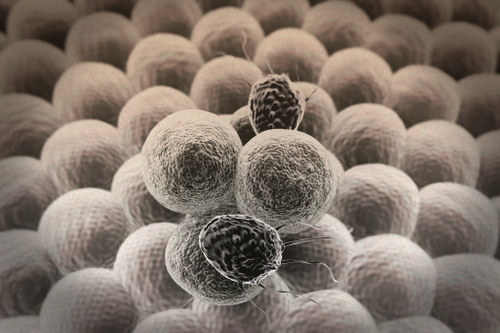
A recent study from the lead scientific advisor on Coronavirus, has found that 82% of those studied with the virus developed Lymphopenia (abnormally low lymphocytes in the blood)
Dr Zhong Nanshan, who discovered the SARS coronavirus in 2003 is the leading advisor in investigating and managing the current coronavirus crisis.
Lymphopenia, which is a condition where a specific white blood cell that is part of the body’s first-line defence against diseases is dramatically reduced.
So, it may be very pertinent to look at the known nutritional elements which nourish the bodies own Lymphocyte making capability, as a means of focus should you feel the need to prepare, eat (or supplement) whilst your body is fighting this Virus.
Here is a strong, concise overview of such dietary elements:
Lymphocyte Nutritional Support:
Dietary Guidelines for a Better Lymphocyte Count You may want to know how to increase lymphocytes naturally. A healthy, nutrient-rich diet can go a long way toward boosting lymphocyte levels. This will provide your immune system with everything it needs to fight off viruses and bacteria that can potentially lead to low lymphocyte levels.
The following is a dietary guideline to follow to help your body improve its lymphocyte count.
Eat lots of lean protein: When the body doesn’t get enough protein, this leads to fewer white blood cells. As a result, you can increase lymphocyte production when you eat protein-rich foods such as grass-fed meats like poultry and beef, organic eggs, wild-caught fish and seafood, and legumes.
Avoid foods high in trans and saturated fats: These fats thicken lymphocytes; as such, reducing trans and saturated fat consumption can help improve immune system health. Avoid unhealthy fats such as margarine, fried foods, hydrogenated oils, and processed baked goods.
Consume healthy fats: Omega-3 fatty acids, on the other hand, will increase your lymphocyte count. Include omega-3 fatty acid foods such as avocado, ground flaxseed, hemp seeds, chia seeds, walnuts, sardines, albacore tuna, white fish, Alaskan salmon, herring, and Atlantic mackerel in your diet.
Eat foods high in beta-carotene: Beta-carotene helps boost lymphocyte production. Foods rich in beta-carotene include carrots, sweet potatoes, butternut squash, romaine lettuce, and spinach.
Eat zinc-rich foods: Zinc is needed to make lymphocytes. It also increases levels of NK cells and T cells, which strengthens your immune system. Foods high in zinc include oysters, asparagus, collard greens, spinach, broccoli, sesame seeds, and pumpkin seeds.
Consume foods high in vitamin C: Vitamin C is known to increase the production of white blood cells such as lymphocytes. Foods high in vitamin C include bell peppers, parsley, kale, oranges, raspberries, tomatoes, and celery.
Eat foods loaded with vitamin D: Not getting enough vitamin D can lower lymphocyte levels and weaken your immune system. Foods rich in vitamin D include organic eggs, raw milk, wild-caught salmon, sardines, mackerel, and tuna.
Eat foods high in vitamin E: Vitamin E supports production of NK cells and B cells. Foods rich in vitamin E include sunflower seeds, almonds, kale, spinach, olives, asparagus, and collard greens.
Eat selenium-rich foods: Selenium helps the body produce more white blood cells. Foods high in selenium include cod, shiitake mushrooms, salmon, tuna, eggs, oats, and broccoli.
Eat more garlic: Garlic is known to boost white blood cell production, which increases the number of NK cells. Purchase fresh, powdered, or dried garlic, and use it in your cooking daily.
Drink more green tea: Green tea compounds can boost immunity by fighting viruses that deplete white blood cells.
Low lymphocyte count, also known as lymphocytopenia, is a cause for concern because when lymphocytes (a type of white blood cell) are low, the body’s ability to repel infections is weakened.
ChooseLife : Of Course, I am always focused on the Acid/Alkaline balancing aspects in life, so this is not to discount the strong evidence that initial infectivity and severity it almost certain to be controlled by the hosts pH, if your tissue at the site of infection is below 7 you are much more likely to become infected, if 6 or lower it has been demonstrated in previous strains that the infectivity is 10x higher, Alkaline eating and supplementation is something I am very focused on for me and my family.
pH-Dependent Entry of Severe Acute Respiratory Syndrome Coronavirus Is Mediated by the Spike Glycoprotein and Enhanced by Dendritic Cell Transfer through DC-SIGN
June 2004
ABSTRACT
The severe acute respiratory syndrome coronavirus (SARS-CoV) synthesizes several putative viral envelope proteins, including the spike (S), membrane (M), and small envelope (E) glycoproteins. Although these proteins likely are essential for viral replication, their specific roles in SARS-CoV entry have not been defined. In this report, we show that the SARS-CoV S glycoprotein mediates viral entry through pH-dependent endocytosis. Further, we define its cellular tropism and demonstrate that virus transmission occurs through cell-mediated transfer by dendritic cells. The S glycoprotein was used successfully to pseudotype replication-defective retroviral and lentiviral vectors that readily infected Vero cells as well as primary pulmonary and renal epithelial cells from human, nonhuman primate, and, to a lesser extent, feline species. The tropism of this reporter virus was similar to that of wild-type, replication-competent SARS-CoV, and binding of purified S to susceptible target cells was demonstrated by flow cytometry. Although myeloid dendritic cells were able to interact with S and to bind virus, these cells could not be infected by SARS-CoV. However, these cells were able to transfer the virus to susceptible target cells through a synapse-like structure. Both cell-mediated infection and direct infection were inhibited by anti-S antisera, indicating that strategies directed toward this gene product are likely to confer a therapeutic benefit for antiviral drugs or the development of a SARS vaccine.
The severe acute respiratory syndrome coronavirus (SARS-CoV) is the likely cause of an acute infectious respiratory disorder identified in highly lethal outbreaks during the past year (10, 18, 21, 32, 40). Infection is characterized by acute flu-like symptoms that progress to a severe febrile respiratory illness with significant mortality. Coronaviruses, comprising a genus of the Coronaviridae family, are enveloped positive-strand RNA viruses. In general, coronaviruses cause respiratory and enteric diseases in humans and domestic animals (15, 20). Two previously known human coronaviruses caused only mild upper respiratory infections (15, 20). In contrast, a highly pathogenic, severe respiratory disease is caused by the SARS-CoV, especially in the elderly (44). Coronaviruses can be divided into three serologically distinct groups (15). Phylogenetically, SARS-CoV is not closely related to any of the three groups (26), though it is most similar to the group II coronaviruses (33, 36).
Although the organization of the SARS-CoV genome is related to that of animal coronaviruses, its genetic sequence is unique, and the structure and function of its gene products are not known. At least 14 open reading frames (ORFs) can be identified in its genome (26, 34, 36). Among these, the replicase/transcriptase genes are located in the 5′ portion of the genome. At its 3′ end, the four major structural proteins (S, M, N, and E) are made through different subgenomic RNAs. Based on comparison to animal coronaviruses, three structural gene products are predicted to be present on the viral envelope: the spike (S), membrane (M), and small envelope (E) proteins (20, 26, 34). The structure of the SARS-CoV envelope differs in some respects from that of other enveloped viruses, such as retroviruses and lentiviruses, many of which contain one viral envelope protein.
Envelope or spike proteins from enveloped viruses have been used to pseudotype retroviral and lentiviral vectors for functional and gene transfer studies (29, 35, 43, 45); however, whether coronavirus glycoproteins could pseudotype these viruses was unknown. Here we report that replication-defective retroviral (Moloney murine leukemia virus) and lentiviral (human immunodeficiency virus type 1 [HIV-1]) vectors can be pseudotyped with the SARS-CoV S protein, and the properties of S related to entry have been defined. Using these pseudoviruses, we were able to determine the relative contributions of SARS-CoV envelope proteins to viral entry and fusion and to examine the roles of these different viral envelope gene products with respect to entry, cell specificity, and potential inhibition of viral replication.
Pertinent Extract:
In contrast, influenza and Ebola viruses are prototypes for viruses that utilize a pH-dependent endocytotic pathway (43). To determine the pathway utilized by the SARS-CoV, the pH dependence of the SARS-CoV S-pseudotyped lentiviral vector was analyzed. Addition of ammonium chloride, which prevents acidification of the endosome, caused a dose-dependent reduction in viral entry (Fig. (Fig.1B,1B, left) at concentrations similar to those described for other pH-dependent viral glycoproteins (3, 11, 43). This effect was also observed with another inhibitor of endosomal acidification, bafilomycin, also in a dose-dependent fashion (Fig. (Fig.1B,1B, right).
Previous research from Meridian Institute Article :
Possible Relevance to SARS
The World Health Organization has concluded that SARS is produced by a new virulent strain of coronavirus. Specific research on the possible pH dependency of the SARS virus has not yet been done. It is well known that coronavirus infectivity is exquisitely sensitive to pH. For example, the MHV-A59 strain of coronavirus is quite stable at pH 6.0 (acidic) but becomes rapidly and irreversibly inactivated by brief treatment at pH 8.0 (alkaline). Human coronavirus strain 229E is maximally infective at pH 6.0. Infection of cells by murine coronavirus A59 at pH 6.0 (acidic) rather than pH 7.0 (neutral) yields a tenfold increase in the infectivity of the virus.
ChooseLife : If the strain of coronavirus responsible for SARS shares the pH characteristics of these other coronaviruses that are pH-dependent, this could be a valuable clue to effective prevention and treatment strategies for this potential epidemic. Perhaps keeping a balanced or slightly alkaline pH environment for the body’s tissues can provide viral protection or enhanced healing for SARS and common viral agents that cause respiratory infections.
Inter-related to this, is research on MUC5B, which has shown that those of lower pH, are much more prone to having inhibited mucous membrane formation:
“Moreover, we demonstrate that the conformation of these highly entangled linear polymers is sensitive to calcium concentration and changes in pH. In the presence of calcium (Ca2+, 10 mM) at pH 5.0, MUC5B adopted a compact conformation which was lost either upon removal of calcium with EGTA, or by increasing the pH to 7.4. These results suggest a pathway of mucin collapse to enable intracellular packaging and mechanisms driving mucin expansion following secretion. They also point to the importance of the tight control of calcium and pHduring different stages of mucin biosynthesis and secretion, and in the generation of correct mucus barrier properties. “
ChooseLife Related Thoughts :
The above shows that there are multiple potential protective methodologies in play, some people may feel a glass of cold water with 1/2 teaspoon of Sodium Bicarbonate every two hours on the first day may be effective (outlined at the bottom of this page), this is one method I would consider myself (Arm and Hammer or Bobs Mill being Aluminium free). Also small Sips of highly Alkaline Milk of Magnesia, every hour, may coat the upper respiratory regions fairly well and rapidly bring up the pH, out of the greater danger zones of lower pH < 6.5 (this is my go to for my kids with sniffles or worse), I would likely do this myself for this situation.
Personally I am going to use this outbreak as a good time to bring my own (and childrens) pH up, using methods as above, plus make some Moreless Alkalising Mineral Mixture, which both Alkalises and significantly raises the Calcium levels in the body but in a complexed form (pre-bonded to Molasses or Honey) which does not hamper the Mucous membrane process outlined above, which as shown above in the scientfic literature is exactly what our bodies need to be ready to either repel, or minimise the effects of such threats.
“The proven value of Bicarbonate of Soda as a therapeutic agent (from a letter to the Church and Dwight Company):
In 1918 and 1919 while fighting the Flu with the U.S. Public Health Service it was brought to my attention that rarely any one who had been thoroughly alkalinized with bicarbonate of soda contracted the disease, and those who did contract it, if alkalinized early, would invariably have mild attacks. I have since that time treated all cases of Cold, Influenza and LaGripe by first giving generous doses of Bicarbonate of Soda, and in many, many instances within 36 hours the symptoms would have entirely abated.
Further, within my own household, before Women’s Clubs and Parent-Teachers’ Association, I have advocated the use of soda as a preventative for ‘Colds’, with the result that now many reports are coming in stating that those who took ‘Soda’ were not affected, while nearly everyone around them had the ‘Flu’.
…An occasional three-day course of the Bicarbonate of Soda increases the alkalinity of the blood, assists elimination and increases the resisting power of the body to all Infectious Diseases…
Whenever taking a bicarbonate solution internally, the soda should be dissolved in cold water. In the event of a threatened attack we recommend the following treatment: During the first day take six doses of half a teaspoon of Bicarbonate of Soda in a glass of cool water, at about two hour intervals.”
In 1984, as an undergraduate at the University of Stirling, Scotland and while carrying out my first piece of independent research, I watched for the first time a fish, a salmon parr, die from acute aluminium toxicity. The whole process took less than forty-eight hours. Within six hours, the fish showed signs of distress and its behaviour changed markedly. It proceeded to seek out the corners of the tank, pushing its head and body against the side of the tank. After twenty-four hours, it began to move randomly and chaotically around the tank before losing its orientation, slipping onto its back, taking a last gasp, before dying. I was left in no doubt about the toxicity of aluminium. I am recalling this event herein because there does seem to be significant complacency concerning the toxicity of aluminium.
An Aluminium Adjuvant is Acutely Toxic Too
In a recent post (https://www.hippocraticpost.com/pharmacy-drugs/the-toxicity-of-aluminium-adjuvants/) I explained why a single injection of a vaccine that includes an aluminium adjuvant is, akin to the salmon above, also an acute exposure to aluminium. It is acute because the total concentration of aluminium in the immediate vicinity of the injection site is extremely high, in the case of a single dose of Infanrix Hexa vaccine, approximately 8000 times higher than is required to kill a salmon parr within forty-eight hours. Even allowing for some dilution of the injected aluminium adjuvant into body fluids bathing and innervating the tissues surrounding the injection site the total concentration of aluminium in a vaccine is sufficient to cause cell death within hours and perhaps minutes of receiving the injection. This is the definition of an acute response, death (cells or whole organism) within a short period of exposure to a toxin. It is a necrotic form of cell death. It initiates an inflammatory response (redness at the injection site). This inflammation drives and perhaps accelerates the subsequent immune response (https://www.hippocraticpost.com/infection-disease/safety-concerns-aluminium-adjuvants/).
A number of mechanisms bring about remediation of acute aluminium toxicity at a vaccine injection site. These are chemical, physical and biological. The toxic free metal ion, Al3+, forms soluble and insoluble complexes with myriad biological molecules while particles of aluminium adjuvant and other insoluble aluminium compounds are taken up by cells infiltrating the vaccine injection site. All of these processes act to reduce the acute toxicity of aluminium at the injection site by lowering the immediate concentration of toxic Al3+. These remedial processes act to secure aluminium in a number of different compartments. All are systemic and all are potential sources of biologically reactive aluminium to the rest of the body. Many chemical compartments where aluminium is bound in myriad different complexes including simple organic moieties like citrate or more complex proteins like the iron transport protein transferrin promote the transport of aluminium away from the injection site.
These processes can be envisaged as continuous passive diffusion of soluble aluminium away from the injection site. The majority of injected aluminium adjuvant is particulate in the first instance and actively taken up, literally eaten, by a number of different cells infiltrating the injection site. Some particles of aluminium adjuvant are taken up by macrophages and thereafter they are retained at or close to the injection site as a granuloma. Generally, these collections of macrophages are considered as benign ‘cancers’ though such descriptions have been coined for situations where the cellular cargo is not aluminium. For example, macrophagic myofasciitis or MMF is a disease, first described by Romain Gherardi in Paris, in which aluminium-rich granulomas at vaccine injection sites are implicated in disease aetiology. Other cells heavily laden with aluminium do not remain close to the injection site and carry their cargo well beyond where the vaccine is administered, for example visiting local lymph nodes as early stops on their travels. Evidence is mounting that these cells may transport aluminium into brain tissue using both lymph and blood as access routes. Perhaps most worrying, evidence of transport of aluminium into brain tissue across the blood-brain barrier and meninges has been shown in autism (https://www.hippocraticpost.com/infection-disease/aluminium-and-autism/).
An Aluminium Adjuvant is a Significant Exposure to Aluminium
With reference to my recent post, an aluminium adjuvant in a vaccine is an acute exposure to aluminium at the vaccine injection site. However, the aluminium content of a single vaccine also represents a significant exposure to aluminium in an infant. For example, the injection of a single dose of Infanrix Hexa into an infant is equivalent to 164 times the daily dose of aluminium in breast milk feeding. Even allowing for an unrealistically high proportion of aluminium being retained in a granuloma at the vaccine injection site (say, for example 40% of the injected aluminium) the daily dose of aluminium in Infanrix Hexa is 100 times higher than an infant receives in breast-feeding. This is a high exposure to aluminium and inevitably results in aluminium being retained in an infant’s tissues, including the infant brain. This is why we must not be complacent about the use of aluminium adjuvants in vaccines. It is why there should be regulations based upon the toxicity of aluminium that govern how much aluminium is allowed in a single vaccine. This limit should be used to give clear unequivocal advice on the number of vaccines that include an aluminium adjuvant that can be given within a specified period. Aluminium is only toxic (as opposed to essential) in the human body and so we should always take every possible step to reduce exposure and ultimately the body burden of aluminium. Infants, due to increased gastrointestinal absorption, reduced urinary excretion and a developing blood-brain-barrier, are uniquely vulnerable to aluminium. We need to protect them from their future now.
Professor in Bioinorganic Chemistry Keele University Honorary Professor, UHI Millennium Institute Group Leader – Bioinorganic Chemistry Laboratory at Keele
Aluminium salts are common adjuvants in vaccines given to children. Their physical, chemical and biological properties have recently been reviewed [1]. However, a debate continues as to whether neonate and infant exposure to aluminium through vaccination is biologically significant with respect to their exposure to aluminium through other routes and especially diet. For example, paediatricians, responsible for administering the vaccine schedule for children, seem in particular, to be uninformed about the properties of aluminium adjuvants and their mode of action in vaccines. This apparent ignorance of the published scientific literature is unexpected in those charged with the wellbeing of neonates and infants and especially in the light of Janeway’s description of alum adjuvant as ‘the immunologist’s dirty little secret’ [2]. Paediatricians such as recently (07/04/2019) Andrew Pollard in The Sunday Times, have a habit of reverting to pure ‘baby talk’ when for example; describing how much aluminium is present in an infant vaccine. They use terms such as ‘minuscule’ and ‘teeny-weeny’ to tell anyone, who asks, how little aluminium there is in a vaccine. They usually then proceed to compare the amount of aluminium in a vaccine with the amount of aluminium in (an adult’s) diet. There are, of course, more accurate, understandable ways to inform parents and other interested parties how much aluminium is present in a vaccine, and I shall endeavour to achieve this herein. An appreciation of how much aluminium is present in a single injection of a vaccine is critical to understanding how aluminium adjuvants are effective in stimulating the immune response.
2. How much aluminium is found in vaccines?
Currently about 20 childhood vaccines include an aluminium adjuvant. Vaccine industry literature (for example; https://www.medicines.org.uk/emc/product/2586/smpc) expresses the aluminium content of an individual vaccine as an amount (weight) of aluminium (not aluminium salt) per unit volume of a vaccine (usually 0.5 mL). Industry does this to account for the fact that there are no strict molecular weights for the polymeric aluminium salts that are used as adjuvants in vaccinations. They prepare acid digests of the adjuvants and measure their total aluminium using ICP MS. This is not explained in the literature they provide with vaccines and can cause confusion for some as the actual weight of hydrated aluminium salt (e.g. aluminium oxyhydroxide, aluminium hydroxyphosphate and aluminium hydroxyphosphatesulphate) in any vaccine preparation is actually approximately ten fold higher. The aluminium salt is the major component of a vaccine (after water) and its high content is why vaccine preparations are invariably cloudy in appearance [1]. As an example, GlaxoSmithKline’s Infanrix Hexa vaccine is reported by the manufacturer to contain 0.82 mg of aluminium per vaccine (0.5 mL). Thus, the weight of aluminium salt in this vaccine is approximately 8 mg, which is approximately ten times the weight of all of the other components of the vaccine when combined. An aluminium-adjuvanted vaccine is essentially a very high concentration of an aluminium salt (8 mg/0.5 mL or 16 mg/mL or 16 g/L) in which just μg of other vaccine components including antigens and other excipients are occluded.
3. Is the amount of aluminium in a vaccine ‘minuscule’?
Generally, in the United Kingdom the first dose of Infanrix Hexa vaccine is injected into muscle when an infant is 8 weeks old. All 8 mg of the aluminium salt (or 0.82 mg of aluminium) will immediately be systemic; it is inside the infant’s body. The repercussions of this being that the injected aluminium may only leave the body through its excretion in either the infant’s urine or sweat. What is the immediate biological response to this exposure to aluminium adjuvant? Aluminium is described as a silent visitor to the human body. What this means is that in the evolution of life on Earth and latterly human evolution, no historic signature is found as evidence for previous exposure to aluminium [3]. By way of comparison with another toxic and non-essential metal, if the adjuvant used in a vaccine was composed of a cadmium salt its injection would immediately initiate a counter-response by the body in an attempt to remove the toxicant. Proteins known to bind and help in the detoxification of cadmium are produced and this is a sure sign that biochemistry had previously encountered non-essential cadmium and selected it out of essential biochemical pathways. Such restorative attempts at detoxification are not triggered for biologically available aluminium and so the ‘processing’ of aluminium adjuvant at the injection site of a vaccine is completely adventitious and one might suggest, random and chaotic. The latter because the fate of aluminium in the body, unlike essential and other non-essential metals, is not subject to any form of homeostasis. Myriad chemical and biological processes will initiate the slow redistribution of the injected aluminium throughout the infant’s body. These steps will involve the processes of disaggregation, dissolution, complexation, precipitation, distribution, cellular uptake and translocation. The description of each one of these processes is an essay in itself and we have addressed them all in many complementary publications [1]. An important and vaccination-specific distinction to make at this point and to carry forward to the following discussion is that aluminium injected into muscle as an adjuvant in a vaccine potentially has uninterrupted access to the infant brain. This is because there is no prerequisite for its passage via the liver, the most prominent organ of detoxification in humans.
We asked if 0.82 mg of systemically available aluminium administered as a single dose in a vaccine is, as some paediatricians would suggest, a minuscule amount of aluminium, for example, as compared to aluminium in the diet. Infants receiving Infanrix Hexa vaccine at 8 weeks of age are concurrently either being breast or formula fed. Data show that the former is likely to result in an 8 week old infant ingesting up to 0.1 mg of aluminium each day [4,5]. On the day an infant receives 8 mg of an aluminium salt, or 0.82 mg of aluminium, in a vaccine it will also ingest 0.1 mg of aluminium in breast milk. However, what proportion of this 0.1 mg of dietary aluminium will be absorbed across the infant gut? Previous research has asked a similar question [6]. The reality is that data for the absorption of aluminium across the infant gut do not presently exist and one has to apply gastrointestinal absorption data obtained for adults. The oft-cited value for adults is that less than 0.1% of ingested aluminium in diet is actually absorbed [7]. The infant gut at 8 weeks is incomplete [8] and is likely to be much more permeable to dietary aluminium, perhaps as much as 100 times more permeable. Applying such clearly conditional criteria it can be estimated that 10% of ingested aluminium or 0.01 mg/day of aluminium in breast milk is absorbed across the infant gastrointestinal tract. However, the blood carrying nutrients and toxins that have been absorbed from the gut, to the rest of the body must first pass through the liver, the major detoxification system of the body. Data on the efficiency of the liver in removing aluminium from the blood is, at best, incomplete in adults [9] and completely unknown in infants. If it is estimated that the liver is 75% efficient in this respect for adults then it is probably only 50% efficient in an infant. When these various conditional factors are accounted for it can be estimated that an infant’s exposure to systemically available aluminium from breast-feeding is approximately 0.005 mg of aluminium each day. In essence during the first 8 weeks or 56 days of life, breast-feeding ostensibly drip feeds an infant with a combined total of 0.28 mg of systemically available aluminium. On day 56 the infant receives a single dose of 0.82 mg of aluminium in the Infanrix Hexa vaccine, a dose equivalent to 3 times the amount of aluminium the infant received during the entire 55 days of life prior to its vaccination. It is well known, if highly unfortunate, that infant formulas are heavily contaminated with aluminium [10,11] and in a worst-case scenario an infant only being formula-fed from birth might be exposed to 0.030 mg of aluminium each day up to vaccination on day 56. Even in this worst-case scenario, the exposure to systemically available aluminium on vaccination day is 25 times higher through the vaccine than through the diet.
4. Acute versus chronic exposure to aluminium
Breast or formula feeding in an infant is a chronic exposure to aluminium. The infant is exposed to a small but continuous supply of systemically available aluminium, aluminium that has the potential to be stored in the infant’s body and excreted from the infant’s body in the urine. Perhaps, at no point during continuous chronic (drip feed) exposure in infancy (0–12 months of age) does the concentration of aluminium in any one physiological compartment increase to bring about overt toxicity. How does dietary exposure to aluminium in infants compare to exposure through vaccination, for example, a single Infanrix Hexa vaccine at 8 weeks of age? The concentration of aluminium (not aluminium salt) in an Infanrix Hexa vaccine upon its injection into muscle is, according to the manufacturer, 0.82 mg/0.5 mL or 1.64 mg/mL or 1.64 g/L or approximately 60 mmol/L. This is the concentration of total systemically available aluminium immediately present at the injection site of the vaccine and available to bring about biological effect. Aluminium adjuvants are not inert depots at the vaccine injection site; they are sources of biologically reactive aluminium [1]. This concentration of total aluminium at the injection site of a vaccine can be put into context by examining the cellular toxicity of aluminium [12] and specifically as identified in recent scientific publications. We can ask the question if we would expect this concentration of aluminium to produce biological effects including cell death at the vaccine injection site. A relevant cell to investigate are lymphocytes and research has demonstrated significant genotoxicity in lymphocytes exposed to only 0.020 mmol/L total aluminium [13]. Similarly, in another study using lymphocytes 0.6 mmol/L total aluminium resulted in significant immunosuppression in both T and B-lymphocytes [14]. Clearly, we would expect profound effects on lymphocytes at the injection site of a vaccine where the total aluminium concentration is 60 mmol/L. Macrophages, a characteristically robust cell, are susceptible to aluminium toxicity demonstrating 50% cell death at a total aluminium concentration of 10 mmol/L [15]. Other more sensitive cell lines would include neuroblastoma where cell viability is reduced by 50% by less than 1 mmol/L total aluminium [16] and similarly for primary hippocampal neurons exposed to only 0.05 mmol/L total aluminium [17]. The concentration of systemically available aluminium immediately present at the injection site of a vaccine is very high in comparison to studies on cell cytotoxicity in the scientific literature. It is an acute exposure to aluminium and it results in significant cytotoxicity including necrotic cell death [1]. The resulting tissue inflammation is the characteristic red mark on the skin at the injection point. This acute toxicity in the immediate vicinity of the injection site underlies the success of aluminium salts as adjuvants in vaccinations [1]. However, while some cells, both present at and infiltrating the injection site, are compromised and especially immediately, other cells act to remedy the situation by taking up aluminium adjuvant into their cytoplasm [18]. This action reduces the concentration of biologically reactive (toxic) aluminium at the injection site and locks away potentially cytotoxic aluminium in intracellular vesicles. Herein may be the real issue linking aluminium adjuvants and severe adverse events following a vaccine. These aluminium-loaded cells remain viable for days, potentially weeks, which means that they can transport their cargo of aluminium anywhere in the body including the infant brain. The recruitment of systemic cells including macrophages to the central nervous system is a widely documented phenomenon [19]. There is now a viable mechanism for the accelerated loading of an infant’s brain with aluminium and evidence to support such a mechanism was demonstrated in our recent paper on aluminium in brain tissue in autism [20].
5. Conclusion: is the amount of aluminium in a vaccine ‘minuscule’?
Simply by looking at just one dose of a vaccine given at 8 weeks of age it is abundantly clear that science does not support this contention, as espoused regularly by many infant paediatricians. In fact, just a single dose of Infanrix Hexa vaccine represents a severe acute exposure to systemically available aluminium. A single dose of this vaccine is equivalent to the exposure to aluminium that an infant would receive from 150 days breast-feeding. It is equivalent to 25 times the daily dose of aluminium received from the most contaminated of infant formulas. It is pertinent to emphasise that an infant would receive a further two doses of this vaccine during the aforementioned 150 day period. It is also highly relevant that other aluminium adjuvanted vaccines, for example Prevenar 13 (https://www.medicines.org.uk/emc/product/453/smpc) and Men B (https://www.medicines.org.uk/emc/product/5168/smpc) are also part of the infant vaccine schedule for this same period. In the United Kingdom it is not uncommon for an infant to receive all three of these aluminium adjuvanted vaccines on the same day. A combined daily exposure of 1.445 mg of aluminium (according to the manufacturer’s data), equivalent to 260 days exposure to aluminium through breast feeding. Exposure to aluminium through a vaccine is, in comparison to diet, an acute exposure and an infant’s physiology will respond differently to exposure to a high concentration of aluminium over a very short time period. The latter, acute versus chronic exposure, while not yet being taken into account in infant vaccination programmes, must now be considered to help to ensure that future vaccination schedules are safe. Currently the EMA and the FDA limit the aluminium content of a vaccine to 1.25 mg (See for example, https://www.ecfr.gov/cgi-bin/text-idx?SID=832c22988b6c802fe810e16ea34ace1a&mc=true&node=se21.7.610_115&rgn=div8). This limit is based upon the aluminium adjuvant’s efficacy in inducing antibody titres. Perhaps now is the time to revise this limit based upon additional factors of vaccine safety.
The effects of calcium ion and broad pH ranges on free fluoride ion aqueous concentrations were measured directly and computed theoretically.
Solubility calculations indicate that blood fluoride concentrations that occur in lethal poisonings would decrease calcium below prevailing levels.
Acute lethal poisoning and also many of the chronic effects of fluoride involve alterations in the chemical activity of calcium by the fluoride ion.
Natural calcium fluoride with low solubility and toxicity from ingestion is distinct from fully soluble toxic industrial fluorides.
The toxicity of fluoride is determined by environmental conditions and the positive cations present. At a pH typical of gastric juice, fluoride is largely protonated as hydrofluoric acid HF.
Industrial fluoride ingested from treated water enters saliva at levels too low to affect dental caries.
Blood levels during lifelong consumption can harm heart, bone, brain, and even developing teeth enamel.
The widespread policy known as water fluoridation is discussed in light of these findings.
Full = https://www.hindawi.com/journals/jeph/2013/439490/