Authors : C. Linhart · D. Davidson · S. Pathmanathan · T. Kamaladas · C. Exley
Abstract:
Human exposure to aluminium is a burgeoning issue. The brain is a sink for systemically available aluminium and a putative target of neurotoxicity. An increasing number of studies continue to confirm the presence of aluminium in human brain tissue though primarily in relation to donors who have died of a neurodegenerative or neurodevelopmental disorder.
Herein, we have measured aluminium in brain tissue in donors who died of a specific disease or condition though without showing any neurodegeneration. The donors were diagnosed as not suffering from multiple sclerosis. Herein, these novel data are compared with recent data on aluminium in brain tissue in multiple sclerosis. Brain tissues from all four lobes were obtained from the Multiple Sclerosis Society Tissue Bank.
Tissues were digested using microwave-assisted acid digestion and their aluminium content was measured by transversely heated graphite furnace atomic absorption spectrometry. Both are established methods in our laboratory. Detailed statistical analyses were used to compare new data with recent data for multiple sclerosis.
Aluminium was found in brain tissue in each donor with a high proportion of measurements (189/291) being below 1.00 μg/g dry weight. The data for all cases (median and IQR) were 0.74 (0.48–1.28), 1.23 (0.62–1.63), 0.84 (0.45–1.14) and 1.01 (0.62–1.65) μg/g dry weight for occipital, parietal, temporal and frontal lobes, respectively.
There was a statistically significant positive correlation between aluminium content of brain tissue and the age of donor. Comparison of data for this non-multiple sclerosis group with brain aluminium data for donors dying with a diagnosis of multiple sclerosis showed that the latter had a statistically significant higher content of brain aluminium.
The data reinforce a previous conclusion that the aluminium content of brain tissue in multiple sclerosis is elevated and support the suggestion that human exposure to aluminium may have a role to play in the aetiology of multiple sclerosis.
New research finds close association between high aluminum content and amyloid-beta
Amsterdam, NL – A new study published in the Journal of Alzheimer’s Disease (JAD) supports a growing body of research that links human exposure to aluminum with Alzheimer’s disease (AD). Researchers found significant amounts of aluminum content in brain tissue from donors with familial AD. The study also found a high degree of co-location with the amyloid-beta protein, which leads to early onset of the disease.
“This is the second study confirming significantly high brain accumulation in familial Alzheimer’s disease, but it is the first to demonstrate an unequivocal association between the location of aluminum and amyloid-beta in the disease. It shows that aluminum and amyloid-beta are intimately woven in the neuropathology,” explained lead investigator Christopher Exley, PhD, Birchall Centre, Lennard-Jones Laboratories, Keele University, Staffordshire, UK.
An association between aluminum and amyloid-beta has been suggested for over 40 years. In an earlier study, brain tissue from donors in the United Kingdom diagnosed with familial AD showed significant accumulations of aluminum. To further understand this relationship, in the current study the researchers measured aluminum in the brain tissue of a cohort of Colombian donors with familial AD who shared a specific mutation. The mutation leads to elevated levels of amyloid-beta, early disease onset, and an aggressive disease etiology. The levels were compared with a control set of brain tissues from donors with no diagnosis of neuropathological disease. They also used aluminum-specific fluorescence microscopy imaging to investigate the relationship between aluminum and amyloid-beta in familial AD.
The results were striking. The aluminum content of the brain tissue from donors with the genetic mutation was universally high, with 42% of tissues having a level considered pathologically significant, and the levels were significantly higher than those in the control set. The imaging studies identified aluminum deposits in all brain tissues studied. They were predominantly co-located with amyloid-beta in senile plaques and occasionally in the brain vasculature. Aluminum was also found separately from amyloid-beta in intracellular compartments including glia and neuronal axons. The results strongly suggest that genetic predispositions known to increase amyloid-beta in brain tissue also predispose individuals to accumulate and retain aluminum in brain tissue.
“Compelling localization of aluminum with a central player in AD, amyloid-beta, strengthens the link of aluminum to the pathogenesis of AD,” commented George Perry, PhD, Professor of Biology, Semmes Distinguished University Chair in Neurobiology, University of Texas at San Antonio, and Editor-in-Chief of JAD.
“One could envisage increased amyloid-beta in brain tissue as a response to high levels of aluminum content, or that aluminum fosters the accumulation of amyloid-beta,” said Dr. Exley. “Either way, the new research confirms my resolve that within the normal lifespan of humans, there would not be any AD if there were no aluminum in the brain tissue. No aluminum, no AD.”
Amyloid-beta (green fluorescence) and aluminum (orange fluorescence) in senile plaque from brain tissue of a familial Alzheimer’s disease donor
In 1984, as an undergraduate at the University of Stirling, Scotland and while carrying out my first piece of independent research, I watched for the first time a fish, a salmon parr, die from acute aluminium toxicity. The whole process took less than forty-eight hours. Within six hours, the fish showed signs of distress and its behaviour changed markedly. It proceeded to seek out the corners of the tank, pushing its head and body against the side of the tank. After twenty-four hours, it began to move randomly and chaotically around the tank before losing its orientation, slipping onto its back, taking a last gasp, before dying. I was left in no doubt about the toxicity of aluminium. I am recalling this event herein because there does seem to be significant complacency concerning the toxicity of aluminium.
An Aluminium Adjuvant is Acutely Toxic Too
In a recent post (https://www.hippocraticpost.com/pharmacy-drugs/the-toxicity-of-aluminium-adjuvants/) I explained why a single injection of a vaccine that includes an aluminium adjuvant is, akin to the salmon above, also an acute exposure to aluminium. It is acute because the total concentration of aluminium in the immediate vicinity of the injection site is extremely high, in the case of a single dose of Infanrix Hexa vaccine, approximately 8000 times higher than is required to kill a salmon parr within forty-eight hours. Even allowing for some dilution of the injected aluminium adjuvant into body fluids bathing and innervating the tissues surrounding the injection site the total concentration of aluminium in a vaccine is sufficient to cause cell death within hours and perhaps minutes of receiving the injection. This is the definition of an acute response, death (cells or whole organism) within a short period of exposure to a toxin. It is a necrotic form of cell death. It initiates an inflammatory response (redness at the injection site). This inflammation drives and perhaps accelerates the subsequent immune response (https://www.hippocraticpost.com/infection-disease/safety-concerns-aluminium-adjuvants/).
A number of mechanisms bring about remediation of acute aluminium toxicity at a vaccine injection site. These are chemical, physical and biological. The toxic free metal ion, Al3+, forms soluble and insoluble complexes with myriad biological molecules while particles of aluminium adjuvant and other insoluble aluminium compounds are taken up by cells infiltrating the vaccine injection site. All of these processes act to reduce the acute toxicity of aluminium at the injection site by lowering the immediate concentration of toxic Al3+. These remedial processes act to secure aluminium in a number of different compartments. All are systemic and all are potential sources of biologically reactive aluminium to the rest of the body. Many chemical compartments where aluminium is bound in myriad different complexes including simple organic moieties like citrate or more complex proteins like the iron transport protein transferrin promote the transport of aluminium away from the injection site.
These processes can be envisaged as continuous passive diffusion of soluble aluminium away from the injection site. The majority of injected aluminium adjuvant is particulate in the first instance and actively taken up, literally eaten, by a number of different cells infiltrating the injection site. Some particles of aluminium adjuvant are taken up by macrophages and thereafter they are retained at or close to the injection site as a granuloma. Generally, these collections of macrophages are considered as benign ‘cancers’ though such descriptions have been coined for situations where the cellular cargo is not aluminium. For example, macrophagic myofasciitis or MMF is a disease, first described by Romain Gherardi in Paris, in which aluminium-rich granulomas at vaccine injection sites are implicated in disease aetiology. Other cells heavily laden with aluminium do not remain close to the injection site and carry their cargo well beyond where the vaccine is administered, for example visiting local lymph nodes as early stops on their travels. Evidence is mounting that these cells may transport aluminium into brain tissue using both lymph and blood as access routes. Perhaps most worrying, evidence of transport of aluminium into brain tissue across the blood-brain barrier and meninges has been shown in autism (https://www.hippocraticpost.com/infection-disease/aluminium-and-autism/).
An Aluminium Adjuvant is a Significant Exposure to Aluminium
With reference to my recent post, an aluminium adjuvant in a vaccine is an acute exposure to aluminium at the vaccine injection site. However, the aluminium content of a single vaccine also represents a significant exposure to aluminium in an infant. For example, the injection of a single dose of Infanrix Hexa into an infant is equivalent to 164 times the daily dose of aluminium in breast milk feeding. Even allowing for an unrealistically high proportion of aluminium being retained in a granuloma at the vaccine injection site (say, for example 40% of the injected aluminium) the daily dose of aluminium in Infanrix Hexa is 100 times higher than an infant receives in breast-feeding. This is a high exposure to aluminium and inevitably results in aluminium being retained in an infant’s tissues, including the infant brain. This is why we must not be complacent about the use of aluminium adjuvants in vaccines. It is why there should be regulations based upon the toxicity of aluminium that govern how much aluminium is allowed in a single vaccine. This limit should be used to give clear unequivocal advice on the number of vaccines that include an aluminium adjuvant that can be given within a specified period. Aluminium is only toxic (as opposed to essential) in the human body and so we should always take every possible step to reduce exposure and ultimately the body burden of aluminium. Infants, due to increased gastrointestinal absorption, reduced urinary excretion and a developing blood-brain-barrier, are uniquely vulnerable to aluminium. We need to protect them from their future now.
Professor in Bioinorganic Chemistry Keele University Honorary Professor, UHI Millennium Institute Group Leader – Bioinorganic Chemistry Laboratory at Keele
Aluminium salts are common adjuvants in vaccines given to children. Their physical, chemical and biological properties have recently been reviewed [1]. However, a debate continues as to whether neonate and infant exposure to aluminium through vaccination is biologically significant with respect to their exposure to aluminium through other routes and especially diet. For example, paediatricians, responsible for administering the vaccine schedule for children, seem in particular, to be uninformed about the properties of aluminium adjuvants and their mode of action in vaccines. This apparent ignorance of the published scientific literature is unexpected in those charged with the wellbeing of neonates and infants and especially in the light of Janeway’s description of alum adjuvant as ‘the immunologist’s dirty little secret’ [2]. Paediatricians such as recently (07/04/2019) Andrew Pollard in The Sunday Times, have a habit of reverting to pure ‘baby talk’ when for example; describing how much aluminium is present in an infant vaccine. They use terms such as ‘minuscule’ and ‘teeny-weeny’ to tell anyone, who asks, how little aluminium there is in a vaccine. They usually then proceed to compare the amount of aluminium in a vaccine with the amount of aluminium in (an adult’s) diet. There are, of course, more accurate, understandable ways to inform parents and other interested parties how much aluminium is present in a vaccine, and I shall endeavour to achieve this herein. An appreciation of how much aluminium is present in a single injection of a vaccine is critical to understanding how aluminium adjuvants are effective in stimulating the immune response.
2. How much aluminium is found in vaccines?
Currently about 20 childhood vaccines include an aluminium adjuvant. Vaccine industry literature (for example; https://www.medicines.org.uk/emc/product/2586/smpc) expresses the aluminium content of an individual vaccine as an amount (weight) of aluminium (not aluminium salt) per unit volume of a vaccine (usually 0.5 mL). Industry does this to account for the fact that there are no strict molecular weights for the polymeric aluminium salts that are used as adjuvants in vaccinations. They prepare acid digests of the adjuvants and measure their total aluminium using ICP MS. This is not explained in the literature they provide with vaccines and can cause confusion for some as the actual weight of hydrated aluminium salt (e.g. aluminium oxyhydroxide, aluminium hydroxyphosphate and aluminium hydroxyphosphatesulphate) in any vaccine preparation is actually approximately ten fold higher. The aluminium salt is the major component of a vaccine (after water) and its high content is why vaccine preparations are invariably cloudy in appearance [1]. As an example, GlaxoSmithKline’s Infanrix Hexa vaccine is reported by the manufacturer to contain 0.82 mg of aluminium per vaccine (0.5 mL). Thus, the weight of aluminium salt in this vaccine is approximately 8 mg, which is approximately ten times the weight of all of the other components of the vaccine when combined. An aluminium-adjuvanted vaccine is essentially a very high concentration of an aluminium salt (8 mg/0.5 mL or 16 mg/mL or 16 g/L) in which just μg of other vaccine components including antigens and other excipients are occluded.
3. Is the amount of aluminium in a vaccine ‘minuscule’?
Generally, in the United Kingdom the first dose of Infanrix Hexa vaccine is injected into muscle when an infant is 8 weeks old. All 8 mg of the aluminium salt (or 0.82 mg of aluminium) will immediately be systemic; it is inside the infant’s body. The repercussions of this being that the injected aluminium may only leave the body through its excretion in either the infant’s urine or sweat. What is the immediate biological response to this exposure to aluminium adjuvant? Aluminium is described as a silent visitor to the human body. What this means is that in the evolution of life on Earth and latterly human evolution, no historic signature is found as evidence for previous exposure to aluminium [3]. By way of comparison with another toxic and non-essential metal, if the adjuvant used in a vaccine was composed of a cadmium salt its injection would immediately initiate a counter-response by the body in an attempt to remove the toxicant. Proteins known to bind and help in the detoxification of cadmium are produced and this is a sure sign that biochemistry had previously encountered non-essential cadmium and selected it out of essential biochemical pathways. Such restorative attempts at detoxification are not triggered for biologically available aluminium and so the ‘processing’ of aluminium adjuvant at the injection site of a vaccine is completely adventitious and one might suggest, random and chaotic. The latter because the fate of aluminium in the body, unlike essential and other non-essential metals, is not subject to any form of homeostasis. Myriad chemical and biological processes will initiate the slow redistribution of the injected aluminium throughout the infant’s body. These steps will involve the processes of disaggregation, dissolution, complexation, precipitation, distribution, cellular uptake and translocation. The description of each one of these processes is an essay in itself and we have addressed them all in many complementary publications [1]. An important and vaccination-specific distinction to make at this point and to carry forward to the following discussion is that aluminium injected into muscle as an adjuvant in a vaccine potentially has uninterrupted access to the infant brain. This is because there is no prerequisite for its passage via the liver, the most prominent organ of detoxification in humans.
We asked if 0.82 mg of systemically available aluminium administered as a single dose in a vaccine is, as some paediatricians would suggest, a minuscule amount of aluminium, for example, as compared to aluminium in the diet. Infants receiving Infanrix Hexa vaccine at 8 weeks of age are concurrently either being breast or formula fed. Data show that the former is likely to result in an 8 week old infant ingesting up to 0.1 mg of aluminium each day [4,5]. On the day an infant receives 8 mg of an aluminium salt, or 0.82 mg of aluminium, in a vaccine it will also ingest 0.1 mg of aluminium in breast milk. However, what proportion of this 0.1 mg of dietary aluminium will be absorbed across the infant gut? Previous research has asked a similar question [6]. The reality is that data for the absorption of aluminium across the infant gut do not presently exist and one has to apply gastrointestinal absorption data obtained for adults. The oft-cited value for adults is that less than 0.1% of ingested aluminium in diet is actually absorbed [7]. The infant gut at 8 weeks is incomplete [8] and is likely to be much more permeable to dietary aluminium, perhaps as much as 100 times more permeable. Applying such clearly conditional criteria it can be estimated that 10% of ingested aluminium or 0.01 mg/day of aluminium in breast milk is absorbed across the infant gastrointestinal tract. However, the blood carrying nutrients and toxins that have been absorbed from the gut, to the rest of the body must first pass through the liver, the major detoxification system of the body. Data on the efficiency of the liver in removing aluminium from the blood is, at best, incomplete in adults [9] and completely unknown in infants. If it is estimated that the liver is 75% efficient in this respect for adults then it is probably only 50% efficient in an infant. When these various conditional factors are accounted for it can be estimated that an infant’s exposure to systemically available aluminium from breast-feeding is approximately 0.005 mg of aluminium each day. In essence during the first 8 weeks or 56 days of life, breast-feeding ostensibly drip feeds an infant with a combined total of 0.28 mg of systemically available aluminium. On day 56 the infant receives a single dose of 0.82 mg of aluminium in the Infanrix Hexa vaccine, a dose equivalent to 3 times the amount of aluminium the infant received during the entire 55 days of life prior to its vaccination. It is well known, if highly unfortunate, that infant formulas are heavily contaminated with aluminium [10,11] and in a worst-case scenario an infant only being formula-fed from birth might be exposed to 0.030 mg of aluminium each day up to vaccination on day 56. Even in this worst-case scenario, the exposure to systemically available aluminium on vaccination day is 25 times higher through the vaccine than through the diet.
4. Acute versus chronic exposure to aluminium
Breast or formula feeding in an infant is a chronic exposure to aluminium. The infant is exposed to a small but continuous supply of systemically available aluminium, aluminium that has the potential to be stored in the infant’s body and excreted from the infant’s body in the urine. Perhaps, at no point during continuous chronic (drip feed) exposure in infancy (0–12 months of age) does the concentration of aluminium in any one physiological compartment increase to bring about overt toxicity. How does dietary exposure to aluminium in infants compare to exposure through vaccination, for example, a single Infanrix Hexa vaccine at 8 weeks of age? The concentration of aluminium (not aluminium salt) in an Infanrix Hexa vaccine upon its injection into muscle is, according to the manufacturer, 0.82 mg/0.5 mL or 1.64 mg/mL or 1.64 g/L or approximately 60 mmol/L. This is the concentration of total systemically available aluminium immediately present at the injection site of the vaccine and available to bring about biological effect. Aluminium adjuvants are not inert depots at the vaccine injection site; they are sources of biologically reactive aluminium [1]. This concentration of total aluminium at the injection site of a vaccine can be put into context by examining the cellular toxicity of aluminium [12] and specifically as identified in recent scientific publications. We can ask the question if we would expect this concentration of aluminium to produce biological effects including cell death at the vaccine injection site. A relevant cell to investigate are lymphocytes and research has demonstrated significant genotoxicity in lymphocytes exposed to only 0.020 mmol/L total aluminium [13]. Similarly, in another study using lymphocytes 0.6 mmol/L total aluminium resulted in significant immunosuppression in both T and B-lymphocytes [14]. Clearly, we would expect profound effects on lymphocytes at the injection site of a vaccine where the total aluminium concentration is 60 mmol/L. Macrophages, a characteristically robust cell, are susceptible to aluminium toxicity demonstrating 50% cell death at a total aluminium concentration of 10 mmol/L [15]. Other more sensitive cell lines would include neuroblastoma where cell viability is reduced by 50% by less than 1 mmol/L total aluminium [16] and similarly for primary hippocampal neurons exposed to only 0.05 mmol/L total aluminium [17]. The concentration of systemically available aluminium immediately present at the injection site of a vaccine is very high in comparison to studies on cell cytotoxicity in the scientific literature. It is an acute exposure to aluminium and it results in significant cytotoxicity including necrotic cell death [1]. The resulting tissue inflammation is the characteristic red mark on the skin at the injection point. This acute toxicity in the immediate vicinity of the injection site underlies the success of aluminium salts as adjuvants in vaccinations [1]. However, while some cells, both present at and infiltrating the injection site, are compromised and especially immediately, other cells act to remedy the situation by taking up aluminium adjuvant into their cytoplasm [18]. This action reduces the concentration of biologically reactive (toxic) aluminium at the injection site and locks away potentially cytotoxic aluminium in intracellular vesicles. Herein may be the real issue linking aluminium adjuvants and severe adverse events following a vaccine. These aluminium-loaded cells remain viable for days, potentially weeks, which means that they can transport their cargo of aluminium anywhere in the body including the infant brain. The recruitment of systemic cells including macrophages to the central nervous system is a widely documented phenomenon [19]. There is now a viable mechanism for the accelerated loading of an infant’s brain with aluminium and evidence to support such a mechanism was demonstrated in our recent paper on aluminium in brain tissue in autism [20].
5. Conclusion: is the amount of aluminium in a vaccine ‘minuscule’?
Simply by looking at just one dose of a vaccine given at 8 weeks of age it is abundantly clear that science does not support this contention, as espoused regularly by many infant paediatricians. In fact, just a single dose of Infanrix Hexa vaccine represents a severe acute exposure to systemically available aluminium. A single dose of this vaccine is equivalent to the exposure to aluminium that an infant would receive from 150 days breast-feeding. It is equivalent to 25 times the daily dose of aluminium received from the most contaminated of infant formulas. It is pertinent to emphasise that an infant would receive a further two doses of this vaccine during the aforementioned 150 day period. It is also highly relevant that other aluminium adjuvanted vaccines, for example Prevenar 13 (https://www.medicines.org.uk/emc/product/453/smpc) and Men B (https://www.medicines.org.uk/emc/product/5168/smpc) are also part of the infant vaccine schedule for this same period. In the United Kingdom it is not uncommon for an infant to receive all three of these aluminium adjuvanted vaccines on the same day. A combined daily exposure of 1.445 mg of aluminium (according to the manufacturer’s data), equivalent to 260 days exposure to aluminium through breast feeding. Exposure to aluminium through a vaccine is, in comparison to diet, an acute exposure and an infant’s physiology will respond differently to exposure to a high concentration of aluminium over a very short time period. The latter, acute versus chronic exposure, while not yet being taken into account in infant vaccination programmes, must now be considered to help to ensure that future vaccination schedules are safe. Currently the EMA and the FDA limit the aluminium content of a vaccine to 1.25 mg (See for example, https://www.ecfr.gov/cgi-bin/text-idx?SID=832c22988b6c802fe810e16ea34ace1a&mc=true&node=se21.7.610_115&rgn=div8). This limit is based upon the aluminium adjuvant’s efficacy in inducing antibody titres. Perhaps now is the time to revise this limit based upon additional factors of vaccine safety.
A burgeoning body of research confirms and affirms the presence of aluminium in human brain tissue. Recently, the first data on aluminium content of brain tissue from donors with diagnoses of familial Alzheimer’s disease, autism spectrum disorder, multiple sclerosis and epilepsy have been published. Quantitative data are supported by aluminium-specific fluorescence microscopy identifying the locations of aluminium in human brain tissue.
The challenge in the future will be to confirm or refute the role played by brain aluminium intoxication in human neurodegenerative disease.
Human exposure to aluminium is burgeoning [1]. Living in the ‘aluminium age’ ensures that the myriad ways in which we are exposed to aluminium today will be even more numerous tomorrow. Aluminium in human brain tissue is the inevitable consequence of this burgeoning exposure [2]. However, what are the consequences of aluminium in human brain tissue? We know that aluminium does not benefit the human brain and in ‘excessive amounts’ it is neurotoxic. The latter label of ‘neurotoxicity’ is widely accepted for humans only because of the experience of relatively rare occurrences of iatrogenic disease [3] and especially dialysis dementia [4]. While aluminium in brain tissue is inevitable when is it also toxic? How can we define toxicity thresholds and when does neurotoxicity achieve a phenotype in affected individuals. How much aluminium in brain tissue is too much?
We reviewed aluminium in human brain tissue almost 10 years ago [2] and in the period since, out with research in our group, there have not been any new quantitative data published on this subject. In 2012, we published the results of our 60 human brain study, an undertaking designed to provide robust and unequivocal data on the aluminium content of human brain tissue [5]. The research applied the most rigorous quality assurance methods possible including the measurement of 174 method blanks. We defined a method blank as an estimate of sample contamination from all experimental procedures beginning with taking tissue samples at the brain bank through to analyses of brain digests by transversely heated graphite furnace atomic absorption spectrometry (TH GFAAS). The only difference with sample treatments being that method blanks did not include brain (any) tissues. The median amount of aluminium present as a contaminant was 22 ng/reaction vessel (used for microwave-assisted digestion) and since the method blank data were not normally distributed, a log transformation was used to determine a contaminant level of 54 ng/reaction vessel (mean + 1.654SD). This value is, thereafter, subtracted from all tissue digests carried out using identical experimental procedures. The final data obtained for 60 human brains were revealing in a number of ways. In the first instance, the median aluminium content for all 713 tissue samples was 1.02 μg/g dry wt., an aluminium content in human brain tissue that might be thought of as inevitable (in donors aged 70–103 years) if not normal. However, scrutiny of data showed that in 41 of the 60 donors, the aluminium content of at least one tissue was above 3.00 μg/g dry wt., an aluminium content thereafter defined as pathologically significant [6]. While the brain bank that provided these tissues refused (retrospectively) to reveal the disease state of the individual donors it is a matter of published record that for this particular brain bank approximately 50% of donors had a diagnosis of dementia at death [7]. As such, the 60 human brain data set for aluminium constituted about 30 donors with a diagnosis of Alzheimer’s disease and 30 aged controls.
The nature of the aluminium data for the 60 human brains, range 0.01–33.00 μg/g dry wt., confirmed that which was already suspected, the focal as opposed to homogenous distribution of aluminium in human brain tissue [8]. It also questioned the value of brain tissue data based solely upon averages. Mean or median values would be informative where tissues were heavily overloaded with aluminium, but generally these statistics are more likely to hide important data on focal accumulations of aluminium. For example, not revealing areas of the brain where the aluminium content is sufficiently high to bring about, at the very least, focal neuropathology. We published the aluminium content of each of the three replicates of each tissue and eschewed the calculation of averages. We have continued the practice of publishing data for all replicates in all subsequent studies on aluminium in human brain tissue.
It is a rare privilege to work with human brain tissue, and weights of tissues available through brain banks are usually limited to less than 1 g. However, on some occasions, an opportunity arises to analyse larger weights of tissue and we had such an opportunity for a case of Alzheimer’s disease linked to occupational exposure to aluminium [9]. We were provided with approximately 20 g frozen weight of the frontal lobe and this was divided into approximately 50 separate samples. Our 60-brain study had identified 0.3–0.5 g as the optimal range for fresh weight of brain tissue. Wet weights below 0.3 g could result in dry weights where the influence of contaminating aluminium might be disproportionately high, whereas wet weights above 0.5 g were prone to incomplete digestion in the microwave. The concentration of aluminium (mean and SD) in the frontal lobe of brain tissue from this occupationally exposed individual was 2.98 (2.73) μg/g dry wt. for 46 replicates. That 35% of the replicates had a concentration of aluminium above 3.00 μg/g dry wt. confirmed a possible role for aluminium in this individual’s early-onset Alzheimer’s disease. Once again, as was the case for brain tissues in the 60-brain study, the range of concentrations was wide from the limit of detection, 0.01–12.97 μg/g dry wt. and demonstrated the focal distribution of aluminium in human brain tissue.
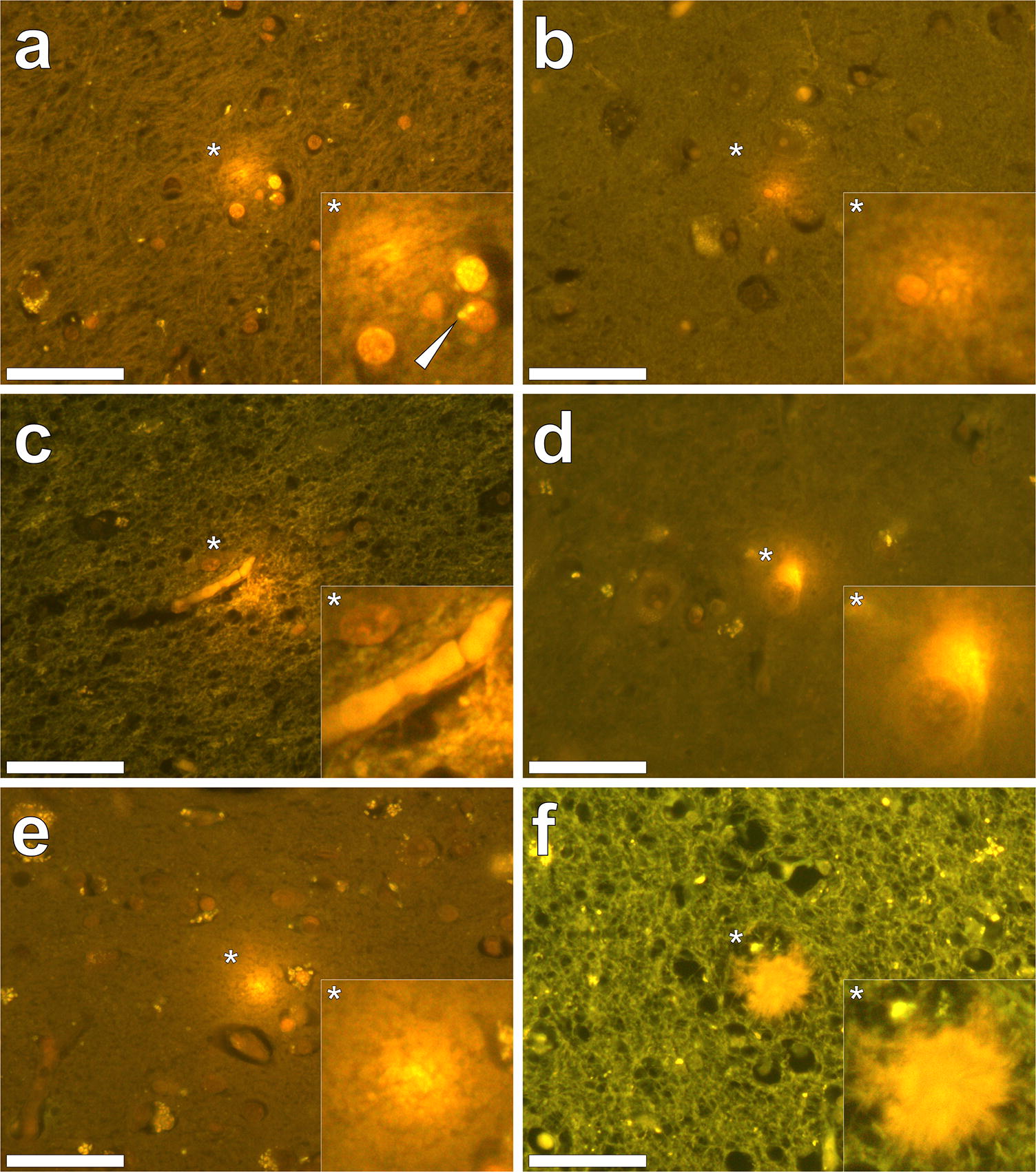
Our subsequent studies have measured the aluminium content of brain tissue from donors who died with a diagnosis of familial Alzheimer’s disease (fAD) [10], autism spectrum disorder (ASD) [11], multiple sclerosis (MS) [12] and epilepsy [13]. In fAD the concentration of aluminium ranged from 0.01 to 35.65 μg/g dry wt. with 11 out of 12 donor brains having at least one tissue where the aluminium content is considered as pathologically significant (≥ 3.00 μg/g dry wt.). For ASD the concentration of aluminium ranged from 0.01 to 22.11 μg/g dry wt. with five out of five donor brains having at least one tissue where the aluminium content was considered as pathologically significant (≥ 3.00 μg/g dry wt.). In MS donor tissue, aluminium concentrations ranged from 0.01 to > 50 μg/g dry wt. and all 14 donors had at least one tissue where the aluminium concentration was considered pathologically significant (≥ 3.00 μg/g dry wt.). Peer review of these published studies has consistently asked how brain aluminium data compare with control donor brains. In addressing this important criticism, a pertinent question is what constitutes control tissue in brain aluminium research. For example, is brain tissue from a donor who has not died from a particular disease, e.g. MS, suitable control tissue for that condition? It should be recognised that aluminium is a toxin and its presence in brain tissue is only inimical. All biologically reactive aluminium will be disruptive and, at the very least, will require energy use in compensating its presence. At some point, compensating a toxic threshold of aluminium will manifest as neurodegeneration identifiable as such by an experienced pathologist. Brain banks are not replete with healthy brains. The majority of brains are from aged donors with underlying conditions. To understand the role of aluminium in neurodegenerative disease including AD, ASD and MS, age- and gender-matched healthy control brains are required. If available, such brain tissues would help in understanding if high concentrations of aluminium contribute towards neurodegeneration, for example, high levels of aluminium with no coincident neurodegeneration would suggest otherwise. Similarly, data on healthy brain tissue would help to identify concentrations of aluminium that would not be expected to contribute towards neurodegeneration. We have worked with a major brain bank in the UK to identify donor brains in which there is little or no neurodegeneration. We have identified 21 brains, the majority from aged donors, and we are currently measuring their aluminium content to help to address the points raised above about what constitutes control brain tissue and what content of aluminium we might expect to find in brain tissue unaffected by overt neurodegenerative disease.Quantitative measurements of brain aluminium are informative, but their interpretation with respect of neurotoxicity requires additional information. Previous research has suggested that the distribution of aluminium in brain tissue is focal, being, for example, associated with neuropathology [2]. The recent development of aluminium-specific fluorescence microscopy [14] has not only been helpful in confirming the presence of aluminium in human brain tissue, but it has also identified the location of aluminium (Fig. 1). Imaging of aluminium has put quantitative data in context, for example, while both fAD and ASD showed some of the highest aluminium content, imaging identified clear differences in the location of aluminium. While in fAD deposits of aluminium were primarily extracellular [10], in ASD aluminium was mainly intracellular and associated with non-neuronal cells such as microglia [11]. Imaging has also helped in identifying associations between aluminium and disease-specific neuropathology. For example, in fAD aluminium was co-localised with amyloid in senile plaques [10], whereas in cerebral amyloid angiopathy (CAA) deposits of aluminium were distinct from those of amyloid [15]. Similarly, while aluminium has been linked historically with neurofibrillary tangles in Alzheimer’s disease [16], tau and aluminium were not co-located in late-onset adult epilepsy [13]. In MS, imaging has identified aluminium in both extra- and intracellular locations as well as associated with neuropathology including plaque-like structures and corpora amylacea [12].
Fig. 1Aluminium imaged in the human brain. Lumogallion staining (1 mM in 50 mM PIPES, pH 7.4, 45 min.) of 5 μm brain tissue sections from donors diagnosed with (a–c) autism spectrum disorder (ASD), d cerebral amyloid angiopathy (CAA), e epilepsy and f familial Alzheimer’s disease (fAD). Intracellular aluminium in astrocytes (a, white arrow), microglia b and in endothelial cells in the lumen of a blood vessel c in donors diagnosed with ASD in a white matter of the hippocampus b the frontal cortex and c white matter of the parietal lobe. Intracellular aluminium in a neuronal cell in a donor diagnosed with CAA in grey matter of the hippocampus (d). Extracellular aluminium in the temporal cortex of a donor diagnosed with epilepsy (e) and (f) co-localised with a senile plaque in white matter of the frontal lobe of a donor diagnosed with fAD. Magnified inserts are denoted with asterisks and images were acquired using an Olympus U-MNIB3 fluorescence filter cube (excitation: 470–495 nm, dichromatic mirror: 505 nm, longpass emission: 510 nm). Magnification ×400, scale bars: 50 μm
Aluminium-specific fluorescence microscopy has confirmed that aluminium is not distributed homogenously in brain tissue and this informs us that even when an aluminium concentration of 1.00 μg/g dry wt. (1 ppm) is determined for 0.500 g fresh weight of tissue, there will be regions within this sample of tissue where the concentration of aluminium is significantly higher than 1 ppm. These total concentrations of aluminium will be sources of biologically reactive aluminium with the potential for toxicity and even cytotoxicity [6]. Local toxicity may result in cellular and neuronal dysfunction and evoke compensatory mechanisms in affected cell populations. Where the concentration of aluminium in 0.500 g of fresh weight of tissue is in excess of 3.00 μg/g dry wt., there will be focal concentrations of aluminium in excess of 10 ppm and the consequent local toxicity will be more overt and compensatory mechanisms less effective. Neurotoxicity is inevitable under such conditions and aluminium will either initiate a disease process or accelerate any disease processes that may be ongoing [17].
The presence of aluminium in brain tissue is an intoxication. It will inevitably exert toxicity at a local level. Future research is required to ascertain the significance of discrete aluminium-related neurotoxicity and how the additive effects of such neuropathologies may eventually contribute towards recognised global neurodegenerative disease characteristic of conditions such as AD, MS, epilepsy and ASD.