Malignancy, on a lovely afternoon tea serving tray, what could possibly go wrong?
In the early 1930’s Otto Warburg won the Nobel Prize.
His work on Cancer Tumour formation, showed that Cells become Anaerobic (run on sugar fermentation), rather than Aerobic (Oxygen) in normal conditions, as a foundational basis of Malignancy.
This was the biggest, most disgustingly ignored, piece of Cancer (any?) research ever. Why?
Sugar and Oxygen share the same space in blood.
So, years after this discovery, Johanna Budwig found that certain long chain fatty acids, and in particular their ”electron cloud bearing characteristics”, were the missing piece to Warburgs puzzle on how Oxygen feeds into the cells, in this dynamic. She should have got a Nobel, as well, of course.
But, she (Budwig) went too far, healing people, or de-activating cancers as she had to label her ideas, due to the Omnipresent Money Men.
So, we know that cells, when they lack about a third oxygen conducting capacity, are in the zone where they may become Cancerous, switch to sugar metabolism when Oxygen is too scarce. Mix emotional anguish in this region = Danger, danger, danger.
So, back to the over-arching point.
MacMillan Cancer have Cake and Coffee mornings, to raise money for Cancer sufferers.
Cake, made from about 20-40% sugar, which will hamper you bloods space to carry oxygen, and 20-40% fats known to strangle the cells Oxygen conductance capacity.
This is madness, this is sickness.
When a friend had Cancer, he disowned me, after I told him of this Sugar/Fat dynamic, his consultant told him my information was ”Unhelpful”, sick, evil, sub-human response.
My dear friend, had a fridge full of Coca-Cola, brought him Kombucha, Sauerkraut, Flaxoil, tried to relay, but he was locked in this ”Fight or Flight” as sugars bore his metabolic burden, he needed to hear from authority he trusted. Instead, his Oncologist turned this help, to anger ”you always think you know best, nobody believes your shit Rich, don’t contact me again”.
When they operated, they told him he nearly died, as the tumour was much bigger than anticipated, go figure, fridge full of Cancer fuel, the Doctors left him feeding from… less support, the Doctors left him feeding from (as they broke our trust with their untruths)…
As, having discussed Warburg with an Oncologist, you see the fear in their eyes, that you know they are serving demons, corporate, demons.
”That’s very old research now”, the Oncologist squirms, like I am giving him a Prostate check, whilst talking.
”Has human physiology changed since then?”, I retort, knowing, of course, it has not.
Then, about a month later, my friend came to this knowledge himself, by then the Oncologist was free of come back, but not his Karma. THEY ALL KNOW.
Where is this place? The arsehole of the world? Want a slice, really gooey, go on, you know you want to…
Shame on you MacMillan, fuelling Cancerous environments, under the guise of helping Cancer.
The upside down modern world, need truth medicine from humans like Budwig and Reams, not corporate parasites selling sickness masquerading as Care.
Let’s shoot those demons down, they’re not here to stay, let’s shoot them down, don’t listen to what they say, make them turn and walk the other way… Nina Kinert, so poetic
Extracted from Dr. Johanna Budwig’s 2005 book “Cancer – The Problem and The Solution”
1998
Lothar Hirneise: What is the fundamental concept of your therapy?
Dr Johanna Budwig: I was senior expert for pharmaceuticals and fats in the Federal Health Office; this was the highest authority in our country responsible for deciding on approvals for medication. At this time, 1951, many applications had been submitted to me for approval, or to be more precise, these were medications for cancer therapy with the sulfhydryl group (sulfur-containing protein compounds). Everywhere I saw that fats played a role, also in expert reports provided by well-known professors like Prof. Nonnenbruch.
Unfortunately, we could only detect fats in the late stage, and there were no chemical values to detect fats chemically at all. By this time, 1951, I had already developed the first chemical verifications for fat, jointly with Professor Kaufmann, the director of the German Federal Institute for Research on Grain, Potatoes and Fat, and my former doctoral advisor, who was also director of the Pharmaceutical Institute. This was published in 1950 in Neue Wege in der Fettforschung (New Directions in Fat Research).
Using the method of paper chromatography, which I had developed, I could analyze 0.1 mg of fat, and characterize it as highly unsaturated or unsaturated. We then published this extensively.
These were the first studies that made it even possible to detect linoleic acid or linolenic acid. Due to the importance of this work, 16 doctoral candidates were assigned to support my efforts. In this situation I noticed the sulfhydryl groups in my appraisal of the medications for cancer therapy. Through official channels I had the right to ask the companies questions relative to how they wanted to substantiate how this substance (sulfhydryl group) could help with cancer. The companies e.g. the company Knoll, which wanted to use these types of medications as cancer therapy were prepared to send all the copies of the created file, on my request.
Consequently, in early 1951, I got a very fast overview of where the problems were in this issue. That was the same year that B. Flaschenträger’s manual appeared: Physiologische Chemie (Physiological Chemistry). The problem of automatic oxygen absorption for the living substrate is one of the most elementary questions in all of physiology, and it is one of the darkest. Everybody knows the sulfur-containing protein compound of the sulfhydryl group can be detected in all breathing tissues.
However another partner must be present in the interplay with this sulfhydryl group, because the self-active oxygen consumption in the living system is executed in a zig zag curve. Strictly speaking, it is manifest that oxygen consumption does not produce a reaction product; rather it occurs in an interplay between the positive electrically-charged sulphur compounds in the protein, and some kind of fatty substance that we cannot detect, because no verifications for it are available. This fatty substance however plays a major role in the Warburg’s respiratory enzyme.
Warburg recognized that with the Warburg respiratory enzyme or the cytochrome oxydase, fats play a role in the lack of oxygen consumption and oxygen utilization (utilization in the living substrate). He wanted to overcome this blockade in the experiment with butyric acid. This attempt was a failure.
Lothar Hirneise: Does this mean that Warburg was the first who attempted to introduce more oxygen into the cell with butyric acid?
Dr Johanna Budwig: No, von Helmholtz […] had attempted to get more oxygen into the cell. He showed that when we treat doves who have become asphyctic (i.e. doves that have been fed in such a manner that oxygen absorption is blocked), with increased ozone or oxygen, they then die more quickly – and this is still the case today. If the “oxygen bomb” is set up in the hospital for a person with oxygen deficiency, then the sick person dies more quickly.
(Chooselife Notes : So, this was known in the 50’s, yet we hooked countless people to respirators during COVID and utterly ignored biological lipid research)
If animals can be made asphyctic through a certain diet, e.g. bleached rice, then they suffocate and neither increased introduction of oxygen nor activation with any other possible substance will help. At this time we already knew vitamin A, B, C, D, and E, but this did not help. Prof. Linus Pauling for example had been involved with animal experiments and knew precisely that it had been published in 1951 that all vitamins had been investigated in searching for the respiratory activator for Warburg’s respiratory enzyme, but this had produced absolutely nothing, not even vitamin C. By the way Professor Pauling, who had requested all my books from me personally, and who also received them from me, never referred to my work later.
(ChooseLife Notes : 2x Nobel Prize winner Pauling, was in the ’Owners Club’ it seems, contextually, little doubt, that Pauling took the FlaxOil blended with Quark after reviewing Budwigs research)
And then the idea occurred to Warburg in 1926 that fats play a role. However he did not know which ones, and experimented with butyric acid. By the way, [Albert] Scent-György also experimented with fats and wrote as early as 1952 that the substances are too easily oxidized and we cannot detect them. In this situation I then published my new ways of fat analysis, namely to introduce sensitive and specific verifications, nice stains. I was able to analyze fats precisely and break them down into the individual fatty acid components.
Lothar Hirneise: What effects do these fatty acids have?
Dr Johanna Budwig: They form the opposite electrical pole to the positively charged protein in the cell nucleus. They are located in the cell membrane and have been known for a long time as lipoids (fatty substances). In the case of tumor formation it was not known why cells in division are present in such great quantities. The wrong concept is still being pursued in medicine today; the tumor is associated with too much growth. This is incorrect. In 1956, I had published that with the tumor many cells in division are present, and amitosis has already started.
However in the case of tumor formation, cleavage of the daughter cell is lacking, which then results in rejection of the aging cells. When a leaf falls from a tree, a skin has formed over it and this vital function for [the normal] growth process is interrupted if the electrically charged fatty acids are lacking.
Lothar Hirneise: How are fatty acids charged electronically?
Dr Johanna Budwig: An important physicist, Kenneth Ford, said in 1966 that the elementary function of the electron exchange with the photons of solar energy is so intense, that one gets the impression the electrical energy stored in the electrons of seed oils, for example, still recognizes the relationship to its forbears in the photons of solar energy. Physically this has been strictly proven. And this absorption of solar energy in seeds is already adjusted in the green leaf to certain very specific wavelengths; science refers to this via the quantosomes. The quanta of physics, of the wave movement are adjusted to the photons of solar energy. This means that they do not even absorb other rays, and if other radiation such as with the transport of nuclear waste (1998) influences human functions, the elementary function between fats and proteins for oxygen absorption, then the radiation from the ashes of the atom bomb work so radically that this dipolarity which must remain in interplay in movement for the life process, is torn apart and the life function is destroyed. In physics these products in physical processes that disturb (i.e. pull apart) the dipolarity in the life process, which really should be banned, have been called radicals for a long time.
Now if you expose a normal life function, regardless of whether it is a seed or a person, to these rays, then intervention in the life function is so radical that in these experiments with quanta biology with the wrong rays even rats and mice jump around until they fall to the floor dead. The term radical comes from physics and was later used as camouflage for the highly heated oils, which we do not want to name by name, but that work exactly the same way.
The base reaction of oxygen consumption and energy extraction from food is executed in the interplay of positive sulfur-containing protein, electronically highly charged with the photons that are suitable in terms of solar energy quanta, stored in seed oils of various quality. This dipolarity differs in quality and it is crucial for the function of the fatty acids that are recognized as vital. Other fatty acids that have a shorter fatty acid chain, e.g. 4, 6, or 8 links, are also utilized under other conditions in the life process.
Here however we are only referring to the fatty acids that are recognized as vital with 18 links in the hydrogen chains (chain with 18 links) and with high electronic enhancement. It is scientifically known that the electronic energy, e.g. in the linoleic fatty acids, is so high-grade that this energy lifts off of the heavy mass and moves as an electron cloud. You must start with the idea that heavy matter, charged positively, pulls down, the electrons pull upwards. In this dipolarity man can stand up, and in this dipolarity man has more capacity to store solar photons than does any other form of life, as demonstrated in the quantum biology of Professor Dessauer in 1954. It is particularly in the brain that these electrons are highly enhanced.
Lothar Hirneise: What is an electron cloud?
Dr Johanna Budwig: If the enhancement of electronic energy is higher through absorption of photons in the electron compounds of the fats, then the power of the electrons is so high in the dipolarity in between gravity and electrons, that they move from the fatty acid (which also contains hydrogen compounds, and which is heavy matter), over the chain with electrons.
Lothar Hirneise: What is the significance of the cloud?
Dr. Johanna Budwig: No life form has as much energy to store the electrons and photons in depots as does man. This electronic energy, stored in the electronic substrate of the human being, stored particularly in the vital, highly unsaturated fatty acids, is such a strong life element for man, that it was correctly said previously that these are certain fatty acids that have been recognized as vital foodstuffs. Man cannot live without them.
For instance, chemists measure the iodine value of fats and say that if oils have a certain iodine value, then these oils are unsaturated. If oils, regardless of whether sunflower oil or flax oil, are treated with overheated steam then these oils can indeed give a positive iodine value, but they are not the vital fats with 18 links, but rather they form cross-links between the fatty acids like a large net, and are highly detrimental to metabolism of the fats, and with protein they act like radicals.
I repeat because it is so important: I have detected particles in oils treated with steam, which indeed have a positive iodine value, but which are highly toxic for man.
Lothar Hirneise: And you attempt to avoid this toxin when treating people who are sick?
Dr. Johanna Budwig In my nutrition therapy I expressly switch off these fats. By the way the discovery of switching-off these toxic oils, has been recognized as nationally worthwhile since 1971. In the oil and fat industry production has not been changed to this day, because they say these products are billion-dollar items, and nobody will give the industry money to establish something new. That is understandable. The representatives of chemotherapy pose another problem. Our chemotherapy is aimed at destruction of the tumor, and it is recognized that chemotherapies destroy many living cells, and the entire person. Anything that disturbs growth is fatal because growth, as elementary life function, is part of the life process of man. We cannot achieve something good with bad tools.
Lothar Hirneise: Can you tell us something more about the unsaturated fatty acids and their net-like connections?
Dr. Johanna Budwig: For a moment try to forget everything that you have read previously. In butter the fatty acids consist of 4 carbon compounds, in coconut fat, goat fat, and sheep fat the fatty acids consist of 6, 8, 10, or 12 carbon compounds respectively. The unsaturated vital fatty acids really start in the chain with 18 carbon compounds. For example, in olive oil there is only one unsaturated bond in the fatty acid. A person who has a deficiency of vital fats does not absorb it. Thus today’s publicity concerning olive oil does not help at all. The fatty acid chains are primarily saturated in the lower area of the carbon chain links, butyric acid, coconut fat, and palm fat, primarily saturated, however they are more easily co-combustible for people if the essential fatty acids are present. The fatty acids with 18 links are the most important essential fatty acids. There are also fatty acids with up to 30 links.
Fatty acids with 18 links, like in sunflower oil or in flax oil with the higher level of unsaturation, are more important for people, particularly for the brain functions of man.
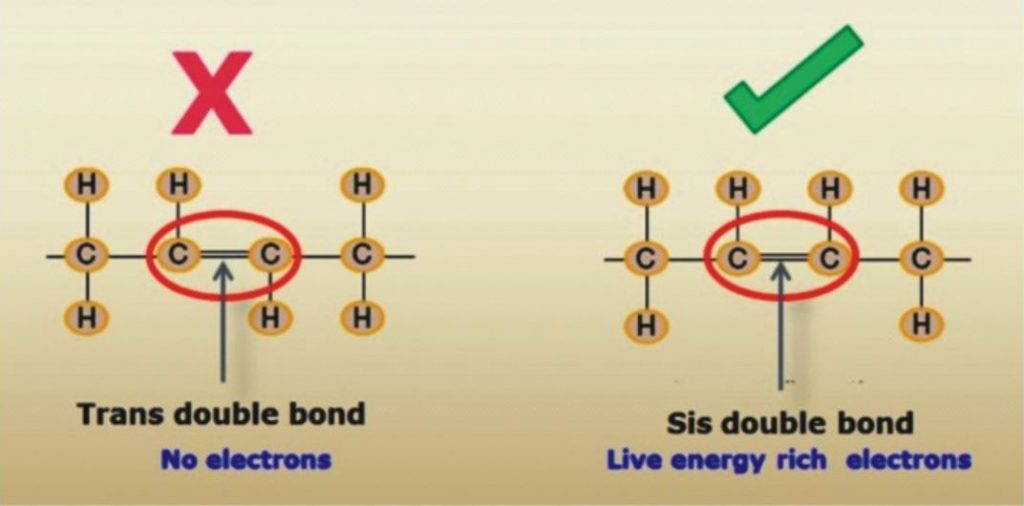
The carbon compounds are heavy matter, even in the fatty acid. If two people reach out to each other with two arms, then they are more strongly bound to each than if they only extend one arm. It is precisely the same for carbon. The fatty acid in olive oil is not considered to be a vital fatty acid; it can only be co-combusted in the organism if essential fatty acids are also present. Linoleic acid rich in electrons is considered vital. There is a particularly high amount of energy in this double double bond of the linoleic acid. This energy wanders and is not fixed in place as it is with a chemical compound, such as with table salt. This energy, wandering between electrons and the positively charged protein with sulfhydryl groups is an alternating association process in the electromagnetic field.This is very important. Perhaps you are familiar with the painting of Michelangelo, where God creates Adam (two fingers pointing to each other, however they do not touch). This is quantum physics, the fingers do not touch. The physicists who I know, Max Planck, or Albert Einstein, or Professor Dessauer all represent the view that man is created by God in His image. You see in being together as human beings there is certainly also a connection without directly touching the other person. People who maintain that they only believe what they can touch are wrong.
The dipolarity with a simple double bond in olive oil is weaker than it is in sunflower seed oil, which is bonded twice. This double double bond is considered to be vital for man. However if the same chain length of 18 carbons has three unsaturated fatty acid compounds, then the electrical energy is as strong as a magnet, depending on the position of the double bond. If double combinations are now closer together, then the energy is greater. All electrical energy that is in movement produces a current that flows and spreads a magnetic field. These electrons also have a magnetic field. You can see this on the window when it rains, for instance. When one drop comes down, it attracts another drop and becomes a larger drop. Precisely the same principle applies with electrons.
This electronic energy is negatively charged. The positively charged sulfhydryl groups of the protein adhere in the unsaturated bonds where the electrons are and that is where they insert their sulphur-containing compounds.
This produces the lipoproteins. The life process is sustained in the interplay between the positively-charged particles and negatively-charged particles. In this process there is no connection, and this is our life element.If a radical break occurs at this point through fatty acids that no longer give off electronic energy, but rather are cross-linked like a net, then the dipolarity can no longer work actively in this net. This is the deadly effect of radicals, because instead of the chains with the electron clouds they interlace a net without electron clouds, indeed with unsaturated bonds, but without dipolarity.
I quickly knew that the triple unsaturated fatty acids, which were called linolenic acid, and which no one had isolated before me, had 18 links and that they did not always carry their double bonds at the same point. They have such a strong electronic energy compared to the heavier matter in the 18-link fatty acid chains, that biologically this energy is far greater than it is with the next arachidon acid with 20 links. The highest electron collection is with the combination of linoleic-linolenic fatty acids in flax oil. The linolenic acid as conjugated (interaction of neighboring double bonds in the molecule that are separated by a single bond) fatty acid is even more effective and is even more strongly effective in interplay with linoleic acid as it is present in the flax oil chain as essential fatty acids for oxygen absorption. This was relatively easy for me to verify in my experiments. I would like to emphasize this. The combination of double unsaturated linoleic acid with triple unsaturated linolenic acid is particularly well-combined in flaxseed. Naturally the quality of flaxseed differs greatly. Naturally red flowering flaxseed and blue flowering flaxseed are naturally different; there is also a difference depending on whether flaxseed is harvested in the cold like in Siberia, or in Africa.
Lothar Hirneise: Is it this energy that heals cancer?
Dr. Johanna Budwig: Yes, this energy is now movable and it is easily released. It is precisely this energy that heals cancer, or does not even allow it to occur. If this vital element is present then no tumor can occur. This vital element is a deciding factor in the immune system. There is a lot of talk these days about immune defense. There is no effective factor in the immune system other than the essential fatty acids. Let’s take for example a patient with a breast tumor. I do not radiate the woman directly on the tumor with my laser, rather I soak the surrounding tissue with fat and protein through the diet, use my oils, e.g. ELDI oils for external application, and radiate the healthy tissue.
Another case. For example I had a child here with a sarcoma in the bone. The child repeatedly claimed that somebody pushed him in kindergarten and he fell on a sharp rock. Since then I have had this. The doctors say, that’s nonsense, falling on a sharp rock does not cause a sarcoma. But I say, if the child’s immune system is weak, and the child is injured, then this can be a trigger.
Lothar Hirneise: But shouldn’t all those people who eat these heated oils get cancer?
Dr. Johanna Budwig: It is very important to view the person as a unit consisting of body, soul, and psyche. However, the factor that is primarily effective at the moment can be quite different. I am still convinced today that if a woman has a very poor marriage, and has to deal with suppression and taunts from her husband day in and day out, then I cannot help her with quark-flax oil. All three factors belong together in a human person. Here is another example. It is very cold here in November. Let’s assume that several people must spend the night outdoors. One takes a hot bath and everything is fine. Another gets lymphogranulomatosis, and another gets pneumonia. They do not all get sick in the same way. The life function of the person always plays a very significant role.
Or let’s look at another case. I have a patient with lung metastases. Her doctor from Ulm came along as well. I counseled her first to change the diet, and then use the oil pack with ELDI oils, to more quickly dissolve the metastasis. The patient was rubbed with oils over the entire body, but naturally in determining the diet and the environment, the whole person must be taken into consideration. This patient knew that she had been given up on. I set up the diet in different steps. Now the patient lives in Ulm and against my advice, she is involved with yoga. In this case I counselled against yoga, and recommended autogenic training, so that her body is pacified and her body is balanced and calm. Sometimes* yoga or sport is very important, but sometimes the patient should not do this.** Guiding the sick person is very important. There is no one treatment that applies for everyone. The patient must also feel for himself what is good for him. Yoga alone cannot replace the healthy basis of nutrition.
Lothar Hirneise: Does this mean that generally you do not recommend physical training [exercise]?
Dr. Johanna Budwig: Correct, it must always be viewed individually. I would never allow a cancer patient with metastases to jog, ride a bicycle, or to practice yoga. His body must relax. I am more an adherent of Zen philosophy in this area. It is based very much on the factor: Let it come. Do not be inactive, rather be very active. I (advise) in my consulting, the patient should not stay in bed, stretch out and let others act. That is incorrect. The patient must work as well. On the other hand, unfortunately I have often experienced what it means when family members do not participate, for example if food is prepared without love, and if the family member basically rejects the treatment. The whole subject of sport must be treated on a very individual basis.
Lothar Hirneise: Do you think that a large tumor must be operated on?
Dr. Johanna Budwig: I can’t make a general statement here. I totally reject radiation and chemo; I also reject hormonal treatment for abdominal cancer. However, operations must be considered very individually. This also applies for tumors in the intestine. I am not a proponent of quickly creating an artificial anus. The ubiquitous technocracy no longer does justice to people.
Lothar Hirneise: How did you get your naturopathic license?
Dr. Johanna Budwig: I acquired my insights in the area of natural science as a well trained expert for fats and pharmaceuticals in a responsible position. Then I tried to make my findings known regarding the significance of the highly unsaturated fats and the damage caused by denatured fats. Naturally this is not necessarily connected with the practice of naturopathy.
However as the number of sick people who sought and received help from me increased, the number of attacks from the medical fraternity who viewed my nutritional advice as an encroachment on their area also increased. Up to 1968, these attacks from doctors were unsuccessful. In 1968, I realized that the use of laser radiation is only possible if the resonance capability is provided in the biological area based on my quantum biological findings. In the USA it was published at that time that the absorption capacity in the living substrate must still be verified.
In this regard, I created the so-called ELDI oils (Electron Differentiation oils) through exact spectroscopic measurements via the absorption of light in different oils. This enabled me to immediately switch the metabolism to positive using the ruby laser that I had chosen. The success was surprising, even to me. These ELDI oils, which are used externally, have a very favorable retuning effect on the metabolism for people who are ill. With my knowledge of the borderline situation concerning the practice of alternative medicine, I said to myself, if I now treat with laser beams, then that constitutes treatment of sick people. Consequently, I obtained the additional permission to practice as a non-medical [naturopathic] practitioner.
Lothar Hirneise: But then you also studied medicine.
Dr. Johanna Budwig: That’s right, in 1955 and in the following years, I also studied medicine, totally real with anatomy and everything that goes along with it. Through the illness of Ms. Martius (wife of the well-known Professor Martius in Göttingen, I am not revealing any unauthorized secret, because this was carried in all the press releases), where I was called on for advice, I was able to use my therapy in various clinics in Göttingen. This was very successful, as is documented in my book Der Tod des Tumors Bd. II (The Death of the Tumor Vol. II). Then, however, I reached a limit as industrial groups and also professors became involved.
Since the people who were opposed to what I was doing always said that I had not studied medicine, I took the time in Göttingen to study medicine. I still remember the time I was working late one night in Göttingen, a woman came to me, with her small child whose arm was supposed to be amputated due to a tumor (sarcoma). I told her what she should do and soon thereafter the subject of amputation was dismissed and the child quickly did very well.
Because I was still a medical student at this time, I was summoned to appear before the Municipal Court due to a petition that I should be prohibited from studying medicine. I was accused of walking through surgery clinics, looking for patients and pulling them out of the clinic. Then I said: “I have never been in the surgery clinic, I do not even know where it is,” I explained that the mother sought me out; I did not seek her out. I then asked what I should do. Shouldn’t I help the child? Because I demonstrably helped a patient successfully (documented in the book Der Tod des Tumors Bd. II (The Death of the Tumor Vol. II), there was a petition to bar me from studying medicine. But the director of the court and counsel for the university, Dr. Henze, rejected this and said: “You have nothing to fear. In my area of jurisdiction nothing will happen to you. If it does there will be a scandal in the scientific community.”
Nevertheless, I concluded that I could better deal with the opponents to my direction as a university graduate and no longer as student. And I still believe today that this decision was right.
Lothar Hirneise: Where have you had the opportunity of presenting your position at congresses and presentations?
Dr. Johanna Budwig: A very important presentation was held in 1964 in the Hilton Hotel in Chicago, at the invitation of the American Oil Chemists Society. Before Professor Kaufmann and I published the validity of my finding concerning the significance of the highly unsaturated fats for the vital function of the human being, Dr. Kaufmann wanted to be sure, and he ordered cytochrome dyed yellow-green in vials from the company Mack in Illertissen. This is considered to be the preliminary stage for hemoglobin, the blood pigment that cancer patients do not produce adequately.
He gave me the yellow-green cytochrome on paper and said: “Touch this and see if it turns red.” I touched it and it turned red, then Prof Kaufmann asked; “Do you have red paint on your finger?” I laughed and said, “No Professor, you can do the same thing as well. Touch it with your fingers.” It turned red, and I said: “I know that you have now started eating flax oil, too.” Then the audience stood up and applauded me. This presentation is in my book Kosmische Kräfte gegen Krebs (Cosmic Powers Against Cancer). Another important presentation was given in Tokyo, where I was the first woman allowed to speak at a congress. At night in my hotel then several women requested that I should give a presentation on the role of women in the world because the newspapers were devoting a lot of attention to the fact that I was the first woman who had been allowed to speak at a conference.
That’s why I was no longer impressed that German doctors who knew too little of real science criticized my work as all philosophy and not science, which is still going on to this day. However I have the impression that the public trend, recognizing the importance of the natural basis for the vital function of the human being, has also become so strong, even in Germany, that pure medical doctrine must join it. The German physician Dr. Roehm who immigrated to the USA published an article in the USA in favor of my work: Who are we, we doctors? In this article he takes the position that it is beneath the dignity of physicians to exclude the natural basis of the life function, as is still attempted in Germany to a great extent. By the way, on the advice of friends in Innsbruck I have put all my scientific work together in a well organized manner (see Presentations at congresses, see Appendix….).
Lothar Hirneise: Does a cell become malignant if it does not get enough oxygen?
Dr. Johanna Budwig: There is an error in your question, because your question reduces the human being to a cell, that is not possible. In humans the lymph system plays a very significant role in the metabolism of fats. When discussing this holistic medical issue you should not localize on the cell, but to return to my previous example – when this child falls on a sharp stone in kindergarten then the important thing is which immune forces are active in the body and which are not.
You see, a doctor comes to me with his 12-year-old child. As I said previously, I always let the sick person speak for himself, this applies to children as well. The boy was happy that he was now allowed [to] speak and said: “OK, now I am going to tell you something. The man who is sitting over there is my father. But the woman sitting next to him is not my mother, that is his secretary, and my mother does not live with us. When my mother lives with us then the muesli tastes good. And when this woman makes the muesli, it doesn’t taste good at all.”
The father turned very red indeed. You see, these things are very important. You cannot reduce the illness to the cells. When people learn that they have been given up on by allopathy then I ask whether the patient is Catholic or Lutheran. And often the patients proceed to tell me for instance how much they are troubled by the fact that they have not been to confession for some time.
Then I tell the patients that before they return in 4 weeks, they should go to confession, and speak openly with their priest. I am Lutheran myself. I always take the whole person very seriously.
Lothar Hirneise: What do you recommend when people do not want to eat quark for ideological reasons or because it does not agree with them?
Dr. Johanna Budwig: A well-known professor from Sweden who is director of a sanatorium for biological therapies, and who knows my treatment very well, called me because he had been called in to treat President Bill Clinton and, according to his information, unfortunately President Clinton cannot eat quark. I did not give him any advice. But I can tell you the following. I have never had a patient who was not able to eat quark in combination with my oil-protein diet.
Lothar Hirneise: What do you recommend to people as a preventative measure so that they don’t get cancer at all?
Dr. Johanna Budwig: Only flax oil as oil. I reject all meat that is in the stores. Fresh meat is OK. Nothing from the frozen food section. You should bake your bread yourself. Oleolux, for example, is something that lasts longer than flaxseed that you can spread on bread or add to vegetables (see the instructions on the next page). You should also prepare your fruit juices yourself. Potatoes are OK, so is cheese.
Also the electromagnetic environment in which we live is very important. Also the textiles that we wear are not insignificant. I reject synthetic materials because they consume too many forces.
I dislike modern foam mattresses because they rob too much energy from you when you sleep. A lot of wood is important in home construction. Carpets are also important so that radiation remains as biological as possible. Gemstones and semi-gem stones are also important because they have good biological radiation and thereby they influence the environment. Books could be written about the favorable effects of gem stones. The environment and living conditions must be as biological [organic & natural] as possible. The lifestyle, e.g. regular sleep is very important. Many cancer patients stay up too late at night and sleep too long in the morning. There is often incorrect or misleading advertising on the labels of our grocery products.
There are hundreds of such aspects that must be taken into consideration. I would never maintain that I heal patients with tumors with the oil-protein diet, on the other hand I get a steady stream of confirmations of my work from recognized experts, that in the surgery clinic in Helsinki they have experienced a success rate of 90% and more, when using the findings of Dr. Budwig, and this success was achieved in cases where allopathy failed. This was submitted by Professor Halme.
Lothar Hirneise: [How] do you work with patients?
Dr. Johanna Budwig: When patients come, I generally schedule two hours for the initial discussion; this is usually between 3:00 and 5:00 in the afternoon. Usually I let the patient report first – about his difficulties, treatments, diagnoses that he is aware of, the medical history, which therapeutic measures have been already employed, etc. In this report, I also learn about the patient’s environment, the workplace, what kind of work the patient has done, and I also learn about the living conditions, e.g. condition of the marriage, and the diet.
Lotus ( Nelumbo nucifera Gaertn.) and Its Bioactive Phytocompounds: A Tribute to Cancer Prevention and Intervention
Abstract
Cancer is one of the major leading causes of death worldwide. Accumulating evidence suggests a strong relationship between specific dietary habits and cancer development. In recent years, a food-based approach for cancer prevention and intervention has been gaining tremendous attention.
Among diverse dietary and medicinal plants, lotus (Nelumbo nucifera Gaertn., family Nymphaeaceae), also known as Indian lotus, sacred lotus or Chinese water lily, has the ability to effectively combat this disease. Various parts of N. nucifera have been utilized as a vegetable as well as an herbal medicine for more than 2000 years in the Asian continent.
The rhizome and seeds of N. nucifera represent the main edible parts. Different parts of N. nucifera have been traditionally used to manage different disorders, such as fever, inflammation, insomnia, nervous disorders, epilepsy, hypertension, cardiovascular diseases, obesity, and hyperlipidemia. It is believed that numerous bioactive components, including alkaloids, polyphenols, terpenoids, steroids, and glycosides, are responsible for its various biological and pharmacological activities, such as antioxidant, anti-inflammatory, immune-modulatory, antiviral, hepatoprotective, cardioprotective, and hypoglycemic activities.
Nevertheless, there is no comprehensive review with an exclusive focus on the anticancer attributes of diverse phytochemicals from different parts of N. nucifera. In this review, we have analyzed the effects of N. nucifera extracts, fractions and pure compounds on various organ-specific cancer cells and tumor models to understand the cancer-preventive and therapeutic potential and underlying cellular and molecular mechanisms of action of this interesting medicinal and dietary plant. In addition, the bioavailability, pharmacokinetics, and possible toxicity of N. nucifera-derived phytochemicals, as well as current limitations, challenges and future research directions, are also presented.
Nutritional Biochemistry and Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama A&M University, Normal, AL 35762, United States.
Abstract
Phytochemicals contribute to the vibrant colors of fruits and it is suggested that the darker the fruit the higher the antioxidative or anticarcinogenic properties. In this study we investigated the possible effects of blueberries (BLU), blackberries (BLK), plums (PLM), mangoes (MAN), pomegranate juice (POJ), watermelon juice (WMJ) and cranberry juice (CBJ) on azoxymethane (AOM)-induced aberrant crypt foci (ACF) in Fisher 344 male rats. Forty-eight male Fisher 344 rats were randomly assigned to eight groups (n=6). The groups were fed AIN-93G as a control (C) diet, the rats fed fruits received AIN-93G+5% fruits and the groups that were given fruits juices received 20% fruit juice instead of water. The rats received subcutaneous injections of AOM at 16 mg/kg body weight at seventh and eighth weeks of age. At 17th week of age, the rats were killed by CO(2) asphyxiation. Total ACF numbers (mean+/-SEM) in the rats fed CON, BLU, BLK, PLM, MNG, POJ, WMJ and CBJ were 171.67+/-5.6, 11.33+/-2.85, 24.0+/-0.58, 33.67+/-0.89, 28.67+/-1.33, 15.67+/-1.86, 24.33+/-3.92 and 39.0+/-15.31. Total glutathione-S-transferase (GST) activity (mICROmol/mg) in the liver of the rats fed fruits (except BLK) and fruit juices were significantly (p<0.05) higher in the rats fed fruits and fruit juices compared with the control. Our findings suggest that among the fruits and fruit juices, BLU and POJ contributed to significant (P<0.05) reductions in the formation of AOM-induced ACF.
The spread of radiofrequency electromagnetic fields (RF-EMF) is rising and health effects are still under investigation. RF-EMF promote oxidative stress, a condition involved in cancer onset, in several acute and chronic diseases and in vascular homeostasis. Although some evidences are still controversial, the WHO IARC classified RF-EMF as “possible carcinogenic to humans”, and more recent studies suggested reproductive, metabolic and neurologic effects of RF-EMF, which are also able to alter bacterial antibiotic resistance. In this evolving scenario, although the biological effects of 5G communication systems are very scarcely investigated, an international action plan for the development of 5G networks has started, with a forthcoming increment in devices and density of small cells, and with the future use of millimeter waves (MMW). Preliminary observations showed that MMW increase skin temperature, alter gene expression, promote cellular proliferation and synthesis of proteins linked with oxidative stress, inflammatory and metabolic processes, could generate ocular damages, affect neuro-muscular dynamics. Further studies are needed to better and independently explore the health effects of RF-EMF in general and of MMW in particular. However, available findings seem sufficient to demonstrate the existence of biomedical effects, to invoke the precautionary principle, to define exposed subjects as potentially vulnerable and to revise existing limits. An adequate knowledge of pathophysiological mechanisms linking RF-EMF exposure to health risk should also be useful in the current clinical practice, in particular in consideration of evidences pointing to extrinsic factors as heavy contributors to cancer risk and to the progressive epidemiological growth of noncommunicable diseases.
Lothar Hirneise: What is your fundamental research?
Dr Johanna Budwig: In 1950, I developed Paper Chromatography of fats. With this technique for first time fats, fatty acids and lipoproteins could be detected directly even in 0.1 ml of blood. Using Co 60 I was successful in producing the first differential reaction for fatty acids, and via radioiodine producing the first direct iodine value. I also developed control of atmosphere in closed system by using gas systems which act as antioxidants. Coloring, separating effects of fats and fatty acids were further developed. Behavior was studied in blue light, red light with fluorescent dyes.
I studied the electrical behavior of the unsaturated fatty acids with their “halo” using dyes with rhodamine red. With this technique I proved that electron rich highly unsaturated Linoleic and Linolenic fatty acids (found abundantly in flax oil) are the undiscovered decisive fats in respiratory enzyme function that Otto Warburg could not find. I studied the electromotoric function of pi-electrons of the linolenic acid in the cell membranes, for all nerve function, secretions, mitosis, as well as cell breakdown. I also examined the synergism of the sulfur containing protein with the Pi-electrons of the highly unsaturated fatty acids and their significance for the formation of the hydrogen bridge between fat and protein, which represent “the only path” for fast and focused transport of electrons during respiration.
This immediately caused a furor. Cancer problem was brought in. Hydrogenated fats, which includes all Trans fatty acids proved to be respiratory poisons. This was extensively published in 1950 in many journals including “New Directions in Fat Research.”
Lothar Hirneise: What is the prime cause of Cancer?
Dr Johanna Budwig: In 1928 Dr. Otto Warburg proved that all normal cells have an absolute requirement for oxygen, but cancer cells can live without oxygen – a rule without exception. Deprive a cell 35% of its oxygen for 48 hours and it may become cancerous. Dr. Otto Warburg has made it clear that the root cause of cancer is oxygen deficiency in the cells, which creates an acidic state in the human body.
He also discovered that cancer cells are anaerobic (do not breathe oxygen), get the energy by fermenting glucose producing lactic acid and cannot survive in the presence of high levels of oxygen. Long back in 1911 Swedish scientist Torsten Thunberg postulated that sulfur containing protein (found in cottage cheese) and some unknown fat is required to attract oxygen in the cell. This fat plays a major role in the cellular respiration. For nearly half century scientists were trying to identify this unknown and mysterious fat but nobody succeeded.
Lothar Hirneise: How did you develop cancer therapy which is called Budwig Protocol?
Dr Johanna Budwig: During my research I found that the blood of seriously ill cancer patients was deficient in these unsaturated omega-3 fats (Linoleic and Linolenic fatty acids), lipoproteins, phosphatides, and hemoglobin. In addition, I noticed that cancer patients had a strange greenish-yellow substance in their blood that I could not find in the blood of healthy people. Then I wanted to develop healing program for cancer.
So I decided to straight waygo for human trials and started to give flax oil and cottage cheese to the cancer patients from four big hospitals in Münster. After just three months, patients began to improve in health and strength, the yellow green substance in their blood began to disappear, tumors gradually receded and at the same time as the nutrients began to rise. Thus I had a cure for cancer. It was a great victory and the first milestone in the battle against cancer. My treatment is based on the consumption of flax seed oil with low fat cottage cheese, raw organic diet, detoxification, mild exercise, flax oil massage and the healing powers of the sun. I have treated approx. 2500 cancer patients during last few decades. Prof. Halme of surgery clinic in Helsinki used to keep records of my patients. According to him my success was over 90% and this was achieved in cases where allopathy failed.
Lothar Hirneise: Can you tell us more about the unsaturated fatty acids and their net-like connections?
Dr. Johanna Budwig: Fatty acid is a carboxylic acid having unbranched chain of 4 to 28 carbons. The saturated fatty acids have primarily short carbon chains. In butter, coconut fat, goat fat and sheep fat the fatty acids consists of 4, 6, 8, 10 or 12 carbons, are saturated, however they are also easily co-combustible if the essential fatty acids are present. The unsaturated vital fatty acids really start with the chain with 18 carbon compounds. There are also fatty acids with up to 30 links. Fatty acids with 18 links, like in flax oil with the higher level of unsaturation, are more important for human, particularly for the brain functions of man. Linoleic acid rich in electrons is considered vital. There is a particularly high amount of energy in this double double bond of the linoleic acid. This energy wanders and is not fixed in place as it is with a chemical compound, such as with table salt. This energy, wandering between electrons and the positively charged protein with sulfur groups is an alternating association process in the electromagnetic field. This is very important. Perhaps you are familiar with the painting of Michelangelo, where God creates Adam (two fingers pointing to each other, however they do not touch). This is quantum physics, the fingers do not touch. The physicists who I know, Max Planck, or Albert Einstein, or Dessauer all represent the view that man is created by God in His image. You see in being together as human beings there is certainly also a connection without directly touching the other person. The dipolarity with a single double bond in olive oil is weaker than it is in sunflower seed oil, which is has two double bonds.
This double double bond is considered to be vital for man. However if the same chain length of 18 carbons has three unsaturated fatty acid compounds, then the electrical energy is as strong as a magnet. This electronic energy is negatively charged. The positively charged sulfur groups of the protein adhere in the unsaturated bonds where the electrons are and that is where they insert their sulfur-containing compounds.
This produces the lipoproteins. The life process is sustained in the interplay between the positively-charged particles and negatively-charged particles. In this process there is no connection, and this is our life element. If radical damage occurs at this point through fatty acids that has lost electron energy, but rather are cross-linked like a net, then the dipolarity can no longer work actively in this net. This is the deadly effect of radicals, because instead of the chains with the electron clouds they interlace a net without electron clouds, indeed with unsaturated bonds, but without dipolarity. I quickly knew that the triple unsaturated fatty acids, which were called linolenic acid, and which no one had isolated before me, had 18 links and that they did not always carry their double bonds at the same point. They have such a strong electronic energy compared to the heavier matter in the 18-link fatty acid chains, that biologically this energy is far greater than it is with the next arachidonic acid with 20 links. The highest electron collection is with the combination of linoleic-linolenic fatty acids in flax oil. The linolenic acid as conjugated (interaction of neighboring double bonds in the molecule that are separated by a single bond) fatty acid is even more effective and is even more strongly interplay with linoleic acid as it is present in the flax oil for oxygen absorption. This was relatively easy for me to verify in my experiments. I would like to emphasize this. The combination of double unsaturated linoleic acid with triple unsaturated linolenic acid is particularly well-combined in flaxseed.
Lothar Hirneise: Is it this energy that heals cancer?
Dr. Johanna Budwig: Yes, this energy is now movable and it is easily released. It is precisely this energy that heals cancer, or does not even allow it to occur. If this vital element is present then no tumor can exist. This vital element is a deciding factor in the immune system. There is no effective factor in the immune system other than the essential fatty acids.
Lothar Hirneise: What is an electron cloud?
Dr Johanna Budwig: If the enhancement of electronic energy is always higher through absorption of sun photons in the unsaturated fatty acids e.g. in linolenic fatty acids, then the power of the electrons is so high in the dipolarity in between gravity and electrons, that they lifts off of the heavy mass and floats like a cloud hence I called them electron cloud.
Lothar Hirneise: What is the significance of the cloud?
Dr. Johanna Budwig: No life form has as much energy to store the electrons and photons as does man. The electronic energy stored particularly in the vital, highly unsaturated fatty acids, is very strong life element for man. Man cannot live without them. If oils are treated with heat and harsh chemicals (during refining and hydrogenation process to increase their shelf life) then the wealth of vital electronic energy is destroyed and Trans fats are formed with net like connections. They are no longer vital fats with 18 links, but rather they form cross-links between the fatty acids like a large net, and are highly damaging to our body, do not adhare with proteins, do not attract oxygen and act like a radicals. I repeat because it is so important: I have detected particles in oils treated with steam, which indeed have a positive iodine value, but which are highly toxic for man.
Lothar Hirneise: So you preach against these toxic hydrogenated and refined oils?
Dr. Johanna Budwig: I am completely against using these “pseudo” fats – “hydrogenated”, “partially hydrogenated”. These are the biggest enemy of mankind. I had scientific proofs. The heart rejects these fats and they are deposited as inorganic fat on the heart muscle itself. They end up blocking circulation, damage heart action, inhibit cell renewal and impede the free flow of blood and lymph fluids.
But it was highly profitable business for multinationals. When I preached against these fats, they stood against me, first they tried to give me hush money and when I refused they filed many fake court cases against me. I was working for humanity and had scientific proof.I was like rock of Gibraltar in my decision; I fought and won all the cases ultimately.
Lothar Hirneise: What is mysterious marriage of sun and moon in terms of quantum physics?
Dr. Johanna Budwig: The photon, the quantum, is the smallest component of the sunlight. It is the fastest moving traveler, it speeds along with time. The photon is always in motion. Nothing can ever halt its motion. The photon is full of all colors, and can change its color and its frequency. The photon is the purest form of energy, the purest wave, can unite with a second photon when it is in resonance with the other, to form a “short-lived particle.” This particle, known as a p0 particle, can break up into two photons again, without mass, as a pure wave in motion. This is the basis for the wonderful back and forth movement between light and matter. The photon can never be pinned down to one location. This is the foundation for the Theory of Relativity.
This very active, dynamic and energetic photon can be captured by electron that are in resonance with the photon. What does that mean? Electrons are already a component of matter. They are also continuously in motion. They constantly oscillate on their own wavelength. They have their own frequency, just like a radio receiver that is set to a specific wavelength.
The electron loves photons. It attracts photons by its magnetic field. When an electrical charge moves, it always produces a magnetic field. The moving photon also has a magnetic field. Both fields attract each other when the wavelengths are in tune. The wave length of the photon – which the photon can change – must fit into the wavelength of the orbiting electron so that the orbit maintains a complete wavelength. This is eternal love between sun-god, photons and moon-goddess, electrons, this is the mysterious marriage of the sun and moon in terms of modern science.This feature is extremely interesting in terms of its physical manifestation, its biological and even its philosophical consequences. All matter has its own inherent vibration. Of course, this also applies to living substances. The energy which is being absorbed must correspond to the inherent wavelength.
The sun rays are very much in harmony with humans. It is no coincidence that we love the sun. The resonance in our biological tissue is so strongly tuned to the absorption of solar energy that there is nothing on earth that has a higher concentration of solar energy photons than humans. This enrichment with solar energy depends strongly on the “like energy” aspects, a wavelength that is compatible with humans, and this is supported when we eat foods that have electrons with electromagnetic fields that attract the photons of the sun. An abundance of these electrons, which are tuned to the solar energy frequency, exist, in many seed-oils. Scientifically these oils have even been designated as electron-rich, “essential,” highly unsaturated fats. But when we began processing fats to increase the shelf life, nobody thought about the significance that this would have for the survival and the further development of the human species. We destroyed their extremely important wealth of electrons, which are very mobile and react so wonderfully to sunlight.
Lothar Hirneise: What is the concept of “human” and “anti-human” in terms of quantum physics?
Dr. Johanna Budwig: It is interesting that in the science of quantum physics the concept of “human” has already been coined. It is “human”, with the highest accumulation of photons, always striving toward the future, who possesses within himself the highest potency of solar energy on the earth. The physicists project from mathematical equations that man, with his wealth of electrons, is directed forwards future in time.
Using mathematical equations that are valid in physics, reversing the time quotient represents the mirror image of man — the “anti-human,” whereas “man” represents the picture of highest rank in terms of physics, i.e. directed against entropy. The “anti-human” is directed back in time. The “anti-human” possesses few solar energy photons, in physical terms electron-poor, directed into the past — also in his thinking — is paralyzed in his life functions, lacks energy and strength because he is missing the electrons that are in harmony with the sun as “life-element.”
The physical processes which are generated through the use of X-rays, gamma rays, atom bombs, or cobalt radiation, are pointed in the same direction as the development toward the “anti-human,” from the perspective of physics and mathematics. The electron structure of the life functions is destroyed by these rays. According to the so-called “World Line” and the Theory of Relativity of modern physics, time and space are connected together in one equation. It is very interesting to investigate our food from this perspective. Fats that have had their electron structure destroyed to make them keep longerpromote the development of the “anti-human”and have a very detrimental effect on the future-directed, electron-rich human being, according to the “World Line diagram.” They disturb the electron exchange within living tissue because they, like tar, act as insulators against electrical conductivity, plainly deaden the life functions at the respective operative locations, e.g. in organs, and in growth centers of the body, as well as throughout the whole body. They also promotes cancer.
On the other hand, electron-rich nutrition, electron-rich highly unsaturated oils,herbs and fruits which are rich in aromatics and natural color components that correspond to the colors of the photon s of sunlight — all these increase the absorption, storage and utilization of the sun’s energy.
Lothar Hirneise: What is your view point about surgery for tumors?
Dr. Johanna Budwig: I am totally against radiation and chemo; I also reject hormonal treatment. Surgery must be considered individually. I am not a proponent of quickly making artificial anus. Conventional oncology no longer does justice to the cancer patients.
Lothar Hirneise: You also studied medicine at the age of 47 years.
Dr. Johanna Budwig: (smiling) Yes handsome! That’s right, my opponents were accusing me that how can I treat cancer patient without doctors degree. This thing pinched me, so in 1955,I joined medical school in Göttingen. There I was using my therapy very successfully in various clinics. I still remember the time I was working late one night in Göttingen, a woman came to me, with her small child whose arm was supposed to be amputated due to a tumor. I treated her and soon the subject of amputation was dismissed and the child quickly did very well.
Because I was still a medical student at this time, I was summoned to appear before the Municipal Court due to a petition that I should be prohibited from studying medicine. I explained the truth in the court. The judge rejected the case and said, “You have done a good job. In my area of jurisdiction nothing will happen to you. If it does there will be a scandal in the scientificcommunity.”
Lothar Hirneise: What do you recommend for prevention of cancer?
Dr. Johanna Budwig: Only flax oil as oil. I reject frozen and preserved meat. Fresh meat is OK. No frozen food and no bakery products. Oleolux should be used as butter. Prepare fruit juices yourself. Cheese and potatoes are OK. Also the electromagnetic environment (e.g. microwave and mobile phones etc.) in which we live is very important. I reject synthetic textiles and foam mattresses because they steal lot of forces from you. A lot of wood in home construction and woolen or silk carpets are also important. Wear gemstones, they also have good biological radiation. The environment and living conditions must be as biological (organic & natural) as possible. Regular sleep is very important.
Authors – Adams LS, Kanaya N, Phung S, Liu Z, Chen S.
Abstract
Previous studies in our laboratory demonstrated that blueberry (BB) extract exhibited antitumor activity against MDA-MB-231 triple negative breast cancer (TNBC) cells and decreased metastatic potential in vitro.
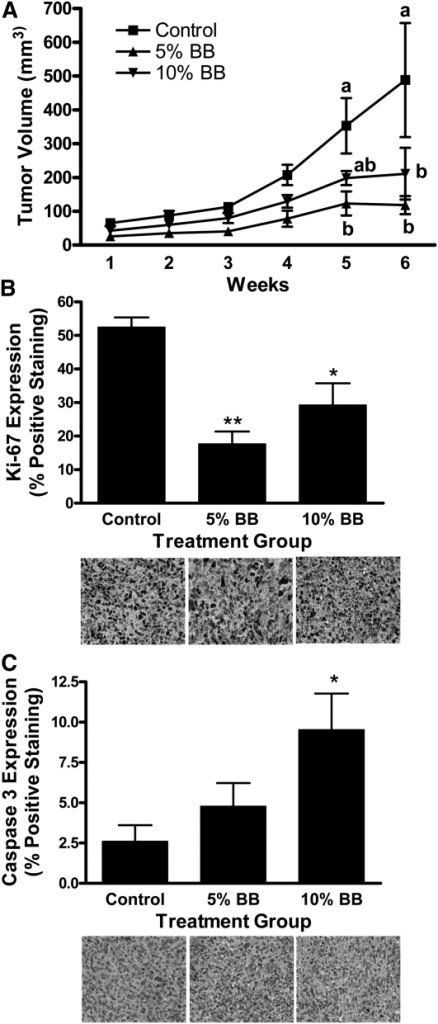
The current study tested 2 doses of whole BB powder, 5 and 10% (wt:wt) in the diet, against MDA-MB-231 tumor growth in female nude mice. In this study, tumor volume was 75% lower in mice fed the 5% BB diet and 60% lower in mice fed the 10% BB diet than in control mice (P ≤ 0.05).
Tumor cell proliferation (Ki-67) was lower in the 5 and 10% BB-fed mice and cell death (Caspase 3) was greater in the 10% BB-fed mice compared to control mice (P ≤ 0.05).
Gene analysis of tumor tissues from the 5% BB-fed mice revealed significantly altered expression of genes important to inflammation, cancer, and metastasis, specifically, Wnt signaling, thrombospondin-2, IL-13, and IFNγ. To confirm effects on Wnt signaling, analysis of tumor tissues from 5% BB-fed mice revealed lower β-catenin expression and glycogen synthase kinase-3β phosphorylation with greater expression of the β-catenin inhibitory protein adenomatous polyposis coli compared to controls.
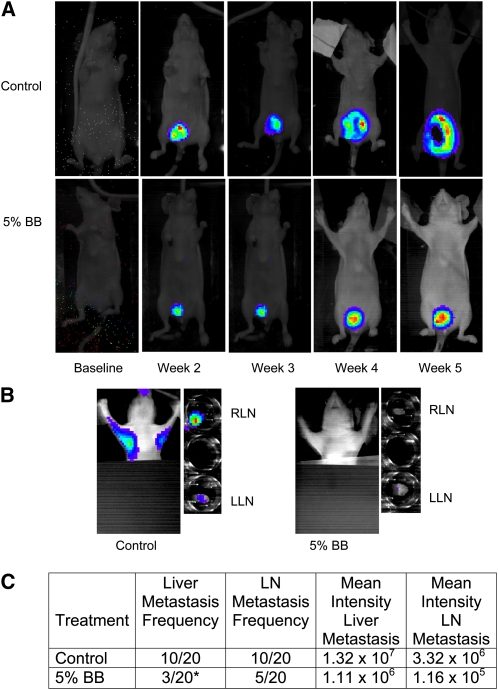
A second study tested the ability of the 5% BB diet to inhibit MDA-MB-231-luc-D3H2LN metastasis in vivo. In this study, 5% BB-fed mice developed 70% fewer liver metastases (P = 0.04) and 25% fewer lymph node metastases (P = 0.09) compared to control mice.
This study demonstrates the oral antitumor and metastasis activity of whole BB powder against TNBC in mice.
MDA-MB-231 tumor volume (A), proliferation (B), and apoptosis (C) in female nude mice fed control, 5% BB powder, or 10% BB powder diet for 8 wk. Data are means ± SEM, n = 6. In A, labeled means at a time without a common letter differ, P ≤ 0.05. In B and C, asterisks indicate different from control: *P ≤ 0.05, **P ≤ 0.01. BB, blueberry. In situ tumor growth and metastasis monitored by Xenogen IVIS imaging at different time points after MDA-MB-231 tumor implantation in female nude mice fed control or 5% BB diet for 7 wk. (A) Orthotopic breast tumor growth from baseline to wk 5 postimplantation. (B) In situ and ex-vivo imaging of RLN and LLN lymph node metastasis at wk 5 postimplantation. (C) Frequency of metastasis and mean intensity of liver and LN metastases analyzed by contingency table. Intensity data are mean ± SEM for the number of mice with metastases. *Different from control, P ≤ 0.05. BB, blueberry; LLN, left lymph node; RLN, right lymph node.
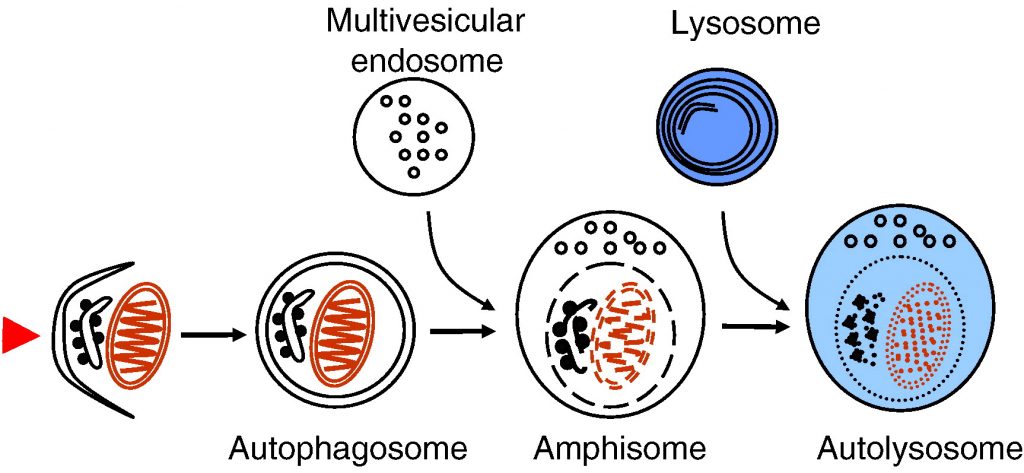
Autophagy delivers cytoplasmic material and organelles to lysosomes for degradation. The formation of autophagosomes is controlled by a specific set of autophagy genes called atg genes. The magnitude of autophagosome formation is tightly regulated by intracellular and extracellular amino acid concentrations and ATP levels via signaling pathways that include the nutrient sensing kinase TOR. Autophagy functions as a stress response that is upregulated by starvation, oxidative stress, or other harmful conditions. Remarkably, autophagy has been shown to possess important housekeeping and quality control functions that contribute to health and longevity. Autophagy plays a role in innate and adaptive immunity, programmed cell death, as well as prevention of cancer, neurodegeneration and aging. In addition, impaired autophagic degradation contributes to the pathogenesis of several human diseases including lysosomal storage disorders and muscle diseases.
Fig. 1. Schematic presentation of autophagosome formation and maturation by fusion with endosomes and lysosomes. The arrowhead on the left illustrates the induction signal that initiates the process.
5. Physiological functions of autophagy
5.1. Stress response and housekeeping function
The role of autophagy as a survival mechanism during short-term amino acid starvation is well documented. Macroautophagy is induced by starvation of serum and amino acids; autophagosomes can be detected already after 15–30 min of starvation [55]. During long-term starvation, chaperone-mediated autophagy increases and macroautophagy decreases [80], [81]. Yeast strains defective in autophagy do not survive nitrogen starvation [38]. Knockout mice deficient in one of the central autophagy proteins, Atg5, show that autophagy is indispensable for the energy metabolism immediately after birth [82]. Atg5 knockout mice die of starvation one day after birth.
In muscle and heart cells, autophagy seems to have a special housekeeping role in the turnover of cytoplasmic constituents including mitochondria. This is revealed by myopathy and cardiomyopathy in patients and mice possessing a defective autophagic degradation due to deficiency of the lysosomal membrane protein LAMP-2 [83], [84], [85]. LAMP-2 deficiency is described in detail later in this review. The importance of autophagy for the heart muscle is supported by a study showing that heart-specific loss of the autophagy protein Atg5 causes cardiomyopathy in mice [86]. Evidence has been published suggesting that damaged mitochondria might be autophagocytosed selectively in a process termed mitophagy [87]. Mitochondria are the major source of reactive oxygen species in cells. Interestingly, reactive oxygen species are necessary for the signal transduction pathway initiating starvation-induced autophagy [88].
It was proposed long ago that autophagy has a role in growth regulation, as suggested by decreased autophagy during growth of the kidney after unilateral nephrectomy [89]. Inducible knockdown of the autophagy protein Atg5 in cell culture shows that autophagy negatively controls cell size [90]. Similar result was observed in Drosophila fat body over-expressing the autophagy protein Atg1 [91].
Autophagy contributes to intracellular quality control and housekeeping, especially in turnover of aggregate-prone proteins. Prevention of autophagy by conditional knockout of atg7 leads to accumulation of ubiquitinated protein aggregates in mouse tissues [92]. Tissue-specific knockout of autophagy proteins in the central nervous system causes accumulation of ubiquitin-positive protein aggregates and neurodegeneration in mice [93], [94]. Further, enhanced autophagy reduces the toxicity of the Huntingtin protein aggregates that accumulate in Huntington disease [95]. Autophagy may prevent aggregate formation by degrading the proteins as monomers, oligomers, or after aggregate formation [96]. It is not clear at present whether aggregated proteins are segregated preferentially, or whether they are removed via unspecific autophagic uptake of cytoplasm. Two proteins have been proposed to function during the uptake of protein aggregates: Alfy and p62 [97], [98]. p62 binds to both ubiquitin-conjugated aggregate-prone proteins and the autophagosome protein LC3 [99], which suggests that it could selectively recruit autophagy machinery to the aggregates and enhance their autophagic clearance.
In addition to removal of cytoplasmic aggregate-prone proteins, autophagy also contributes to the quality control in the ER. Unfolded protein response induces autophagy, and this induction is beneficial for cell survival [100], [101], [102].
Autophagic degradation is also needed for early embryonic development. A recent study shows that autophagy-defective mouse eggs fertilized with autophagy-defective sperm, failed to develop beyond the four and eight cell stages [103]. The authors suggest that autophagy may be needed in the preimplantation embryos for protein recycling, production of amino acids for protein synthesis or substrates for energy production, or for removal of obsolete maternal factors.
5.2. Innate and adaptive immunity
Autophagy contributes to both innate and adaptive immunity [104], [105]. In some cases autophagy can protect cells against intracellular pathogens. Induction of autophagy during Herpes simplex virus infection, and localization of viral particles inside autophagic vacuoles, were proposed to indicate that autophagy acts as a host-defense mechanism in infected cells [59]. The Herpes virus virulence protein, ICP34.5, inhibits autophagy, suggesting that the virus has developed a way to prevent the autophagic defense of the host cell. Autophagy may also help cells to defend against intracellular bacteria [106]. Sequestration of intracellular Group A Streptococci in autophagosome-like structures protects cells against the bacteria [107]. Mycobacterium tuberculosis is normally able to survive inside macrophages by preventing the fusion of phagosomes with lysosomes. Surprisingly, induction of autophagy bypasses the maturation defect, leading to phagolysosome formation and bacterial killing [108].
Macroautophagy also contributes to antigen presentation. Major histocompatibility complex (MHC) class II molecules present products of lysosomal proteolysis to CD4(+) T cells. Extracellular antigen uptake is considered to be the main source of MHC class II ligands. However, it was demonstrated that in MHC class II-positive cells, including dendritic cells, B cells, and epithelial cells, autophagosomes continuously fuse with multivesicular MHC class II-loading compartments [109]. This pathway is of functional relevance, because targeting of the influenza matrix protein 1 to autophagosomes enhances its MHC class II presentation to CD4(+) T cells. Thus it seems that macroautophagy efficiently delivers cytosolic proteins for MHC class II presentation and can improve helper T cell stimulation.
5.3. Cell death
Autophagy also seems to have roles in programmed cell death [110], [111]. Type II programmed cell death, or autophagic cell death, was originally described in mammary carcinoma cells [112], [113]. Autophagy proteins were shown to be necessary for cell death under certain conditions, such as in apoptosis-defective cells [114], [115], [116]. In this scenario autophagy is needed for the execution of cell death. Under other conditions, such as nutrient starvation, autophagy protects cells against apoptosis by providing nutrients [117], [118], [119]. The regulation of apoptosis and autophagy are linked via the antiapoptotic protein Bcl-2. Bcl-2 inhibits Beclin 1-dependent autophagy by binding to Beclin 1 and preventing its association with Vps34 [52]. This anti-autophagy function of Bcl-2 was proposed to help maintain autophagy at levels that are compatible with cell survival, rather than cell death.
Lipids may also regulate autophagy and its outcome to the host cell. Ceramide and sphingosine 1-phosphate, a metabolite of ceramide, both induce autophagy in mammalian cells [120]. The outcome on cell survival is however different: ceramide promotes cell death, while sphingosine 1-phosphate increases cell survival. Ceramide is part of the signaling cascade initiated by chemotherapy, while sphingosine 1-phosphate is part of the signaling cascade initiated by starvation. Beclin 1 level and the autophagy response are stronger during ceramide signaling [121].
Autophagy has functions in cell death during development [122]. atg genes are necessary for the clearance of apoptotic cells during embryonic development in mice [123]. Autophagy contributes to dead-cell clearance during programmed cell death by maintaining cellular energy levels in the dying cells, thereby allowing the generation of cell surface and secreted signals that then promote engulfment of cell corpses by neighboring cells. Autophagy is also indispensable for the execution of certain types of cell death during development. The degradation of Drosophila salivary glands by type II programmed cell death depends on autophagy [73].
5.4. Aging and longevity
Finally, autophagy also contributes to longevity [124]. Reduced caloric intake increases longevity in several animal species. Increased autophagic turnover of cytoplasmic constituents including mitochondria was shown to contribute to the longer life in the dieting animals [125]. Further evidence that autophagy contributes to longevity come from Caenorhabditis elegans mutants possessing a defective insulin receptor (daf2 mutant), which live longer than control worms. The increased lifetime of these mutant worms depends on a functional autophagic pathway [126]. Moreover, knockdown of autophagy gene products including Atg7 and Atg12 were shown to shorten the lifespan of both wild type and daf2 mutant C. elegans[127]. Further, promoting basal levels of autophagy in the nervous system of adult Drosophila enhances longevity of the flies [128]. Together these studies give strong support for a role of autophagy in the prevention of aging.
Fig. 6 summarizes the physiological functions of autophagy described above.
Fig. 6. A summary of the functions of autophagy in health and disease.
6. Autophagy and disease
6.1. Cancer
Impaired autophagy contributes to cancer development [2], [129], [130]. Beclin 1 is monoallelically deleted in a large proportion of human breast and ovarian cancers. Over-expression of Beclin 1 in a breast cancer cell line increases autophagy and decreases the growth and tumorigenicity of these cells [131]. Mice with heterozygous deletion of Beclin 1 have less autophagy and more tumors than control mice [132], [133]. Further, the other autophagy-promoting components of the Beclin 1/Vps34 complex, UVRAG and Ambra 1 (Fig. 4), are also tumor suppressors [48], [49]. Moreover, knockout of Bif-1, also part of the Beclin 1 complex, significantly enhances the development of spontaneous tumors in mice [50]. On the other hand, binding of the proto-oncogenic proteins Bcl-2 or Bcl-XL to Beclin 1 inhibit autophagy [51], [52]. In addition to the Beclin 1 complex, other tumor suppressors also enhance autophagy. PTEN is a phosphatase that decreases the concentration of class I PI3 kinase product and enhances autophagy [72]. PTEN is also a tumor suppressor [134]. Further, the activities of Ras and class I PI3-kinases inhibit autophagy and promote cell growth. Ras is mutated and class I PI3 kinases are upregulated in many cancers [135], [136].
The results described above show that autophagy contributes to the prevention of tumorigenesis. Impaired autophagy can contribute to tumor formation via impaired regulation of cell growth, and/or via decreased cell death. In addition, it was shown that failure to sustain metabolism via autophagy results in increased DNA damage. This chromosomal instability increases tumor progression [137].
In advanced cancers, autophagy may have the opposite effect on the tumor development. Autophagy can benefit the progression of the tumor because it can provide nutrients during starvation [129], [130], [138]. In addition, autophagy was recently shown to improve the survival of p53-deficient cancer cells under starvation or hypoxic conditions [139]. These findings suggest that autophagy inhibition, rather than stimulation, might be beneficial in treatment of advanced cancer.
6.2. Neurodegeneration
Many age-related neurodegenerative diseases are characterized by the accumulation of ubiquitin-positive protein aggregates in affected brain regions. These misfolded, aberrant proteins can disrupt neuronal function and cause neurodegeneration. As described earlier, autophagy is necessary for the clearance of aggregate-prone proteins that are toxic especially for post-mitotic cells like neurons [130]. Tissue-specific knockout of the autophagy genes in neurons causes a massive accumulation of ubiquitin-positive protein aggregates and neurodegeneration in mice [93], [94], indicating that autophagy is needed for the constitutive clearance of aggregate-prone proteins. Autophagy was recently shown to enhance the clearance of Huntingtin, mutant tau, synphilin 1 and α-synuclein, but not AIMP2 (p38) and mutant desmin [140]. This study indicates that autophagy is not able to degrade all protein aggregates. However, the role of autophagy has been demonstrated in Huntington’s disease, caused by mutations in Huntingtin, and familial Parkinson’s disease, caused by mutations in α-synuclein. Enhanced autophagy in animal models of these diseases improves clearance of the aggregated proteins and reduces the symptoms of neurodegeneration [95], [141].
ESCRT complexes are necessary for the biogenesis of multivesicular endosomes. As described earlier, multivesicular endosomes are necessary for the maturation of autophagosomes into degradative autolysosomes. Mutations in ESCRT III subunits CHMP2B or mSnf7-2 are associated with two neurodegenerative diseases, frontotemporal dementia and amyotropic lateral sclerosis. Both diseases are characterized by abnormal ubiquitin-positive protein deposits in affected neurons. Cell lines and fruit flies depleted of CHMP2B or mSnf7-2 show decreased autophagic degradation, increased levels of ubiquitin-positive aggregates and increased neurodegeneration [27], [29].
Alzheimer’s disease is characterized by the accumulation of extracellular amyloid plaques in the brain. These plaques consist of aggregated β-amyloid (Aβ) peptide. Autophagy was proposed to contribute to the production of Aβ. Autophagic compartments containing both amyloid precursor protein and Aβ accumulate in dystrophic neurons in Alzheimer brain [142], [143]. Purified autophagic vacuoles contain all necessary constituents for Aβ production [142], and autophagic compartments were identified as a major reservoir of intracellular Aβ in the brain of Alzheimer patients and mouse models. The primary cause for the increased accumulation of autophagic compartments in Alzheimer’s disease was recently suggested to be their retarded maturation to autolysosomes [144].
A recent study, however, challenges the idea that autophagy contributes to the pathogenesis of Alzheimer’s disease. Beclin 1 was shown to be decreased in affected brain regions of patients with Alzheimer disease early in the disease process [145]. Heterozygous deletion of Beclin 1 in mice decreased neuronal autophagy and resulted in neurodegeneration. Transgenic mice expressing human amyloid precursor protein have been used as a mouse model for this disease. Genetic reduction of Beclin 1 expression increased intraneuronal Aβ accumulation, extracellular Aβ deposition, and neurodegeneration [145]. Increasing Beclin 1 levels by lentiviral expression reduced both intracellular and extracellular amyloid pathology in these transgenic mice. This study suggests that decreased, not increased, autophagy promotes Alzheimer’s disease progression. Further, enhancing autophagy by increasing Beclin 1 levels may have therapeutic potential in this disease.
6.3. Autophagy and lysosomal storage diseases
Niemann–Pick type C is a neurodegenerative lipid storage disorder characterized by a disruption of sphingolipid and cholesterol trafficking caused by mutations in either of two genes, npc1 and npc2. The disease produces cognitive impairment, ataxia and death, often in childhood. Cells deficient in npc genes show increased expression of Beclin 1 and LC3-II, the autophagosome-specific form of LC3, suggesting autophagy is induced [146]. Increased levels of LC3-II have also been observed in npc-deficient brain tissue [147]. npc-Deficient cerebellar Purkinje neurons undergo a cell death that was proposed to depend on autophagy [148], suggesting increased autophagy may be harmful for neurons in NPC patients.
Most lysosomal storage diseases are caused by deficiencies of lysosomal hydrolases, leading to accumulation of undegraded substrate and other material in the lysosomal compartment. Lysosomal accumulation of substrates can also affect autophagosome–lysosome fusion. Autophagosomes accumulate in brain and isolated cell lines of mouse models of two lysosomal storage diseases associated with severe neurodegeneration, multiple sulfatase deficiency and mucopolysaccharidosis type IIIA [149]. Significantly reduced colocalization of the lysosomal membrane protein LAMP-1 with the autophagosome marker LC3 indicates that the fusion of lysosomal compartments with autophagosomes is impaired. In addition, cell lines isolated from these mice have decreased ability to degrade aggregate-prone proteins and show accumulation of polyubiquitinated proteins and non-functional mitochondria. Thus, neurodegeneration observed in many lysosomal storage diseases may be at least partially due to impaired autophagic degradation, which is particularly vital for neurons.
6.4. Autophagy and muscle disorders
Autophagic vacuoles are a frequent feature in numerous muscular disorders. Such a pathological situation can be observed in patients suffering from Danon disease, an inherited disease resulting from null mutations in the lysosomal membrane protein LAMP-2 [83]. LAMP-2 deficiency leads to a fatal cardiomyopathy and myopathy sometimes associated with mental retardation [150]. Accumulation of autophagic vacuoles in the heart and skeletal muscle are hallmarks of the disease [84]. Studies in LAMP-2 deficient mice revealed in part similar findings [85]. Fifty percent of these mice die at an early postnatal age with massive accumulation of autophagic vacuoles in several tissues including liver, pancreas, spleen, kidney, lymph nodes, neutrophilic leukocytes, skeletal muscle, and heart. Autophagic vacuoles containing single mitochondria were frequently observed in cardiomyocytes [151] indicating that mitochondria are a main target for autophagic degradation in muscle tissues. These cellular alterations lead to a reduced contractility and an increased size of the heart in LAMP-2 knockout mice. This is in agreement with the finding that cardiomyopathy is the hallmark in Danon disease patients [83]. Biochemical and electron microscopy studies reveal that a retarded consumption, rather than increased formation, of autophagic vacuoles leads to their accumulation [152]. LAMP-double deficient fibroblasts lack both LAMP-2 and the structurally related LAMP-1 protein. These cells show a defect in the final maturation steps of late autophagic vacuoles, involving retarded fusion with lysosomes [33], [119]. Interestingly, recruitment of the small GTPase Rab7 to autophagosomes is retarded in these cells [32].
Inhibition of lysosomal fusion using hydroxy-chloroquine causes similar vacuolar alterations and myopathies to Danon disease, confirming the important role of lysosome–autophagosome fusion for muscle cell physiology [153]. Impaired autophagosome maturation may also be related to other types of diseases such as X-linked myopathy with excessive autophagy, infantile autophagic vacuolar myopathy, adult-onset autophagic vacuolar myopathy with multiorgan involvement, and X-linked congenital autophagic vacuolar myopathy [154]. The molecular defects in these disorders are still unknown.
Although altered autophagy has been observed in various heart diseases, including cardiac hypertrophy and heart failure, it remains unclear whether autophagy plays a beneficial or detrimental role in these diseases. As mentioned earlier, tissue-specific deletion of atg5 in heart causes cardiac hypertrophy and contractile dysfunction [86]. In addition, increased levels of ubiquitinated proteins and abnormal mitochondria are found, especially after treatment with pressure overload or β-adrenergic stress. This suggests that autophagy is needed in the heart to ensure the availability of sufficient energy substrates and to control cardiomyocyte size and global cardiac structure and function.
6.5. Common aspects in autophagy and phagocytosis
As described above, autophagy plays a role in innate immunity against intracellular pathogens [104], [105] by clearing microbes directly via ingestion into autophagosomes for subsequent degradation in autolysosomes [108], [155]. Similar to the process of intracellular defense, phagocytosis is an evolutionary conserved mechanism involved in the removal of extracellular organisms. Interestingly, it was found that the phagocytic and autophagic pathways are linked. Toll like receptor (TLR) activation triggers the recruitment of autophagy proteins LC3, Atg5 and Atg7 to the phagosomal pathway. Before these events Beclin 1 and class III PI3-kinase activity are found in phagosomes [156]. These autophagy-specific proteins are recruited to the phagosome, while almost no classical autophagosomes are observed in the cells. Phagosome fusion with lysosomes is then initiated, leading to acidification and killing of the ingested organisms. Thus, engaging the autophagy pathway via TLR signaling (especially TLR7 through its binding to single-stranded RNA) enhances phagosome maturation and destruction of pathogens [157]. This underscores the intimate link between autophagy and phagocytosis.
As mentioned earlier, M. tuberculosis is able to survive inside macrophages by preventing the fusion of phagosomes with lysosomes, but induction of autophagy bypasses the maturation defect, leading to phagolysosome formation and bacterial killing [108]. Autophagy induction induces the localization of Beclin 1 and LC3 to phagosomes, suggesting the phagosomes are diverted to an autophagosome-like compartment that is then able to fuse with lysosomes.
The association between autophagy and phagocytosis is also underlined by studies with cells lacking either one or both LAMPs [33]. As described above, autophagosome–lysosome fusion is impaired in LAMP-double deficient cells [32]. Whereas macrophages and fibroblasts from LAMP-1 or LAMP-2 single-deficient mice display normal fusion of lysosomes with phagosomes, in LAMP-double knockout fibroblasts phagosomes are unable to recruit late endosomal/lysosomal markers and phagocytosis is arrested prior to the acquisition of Rab7 [158]. Interestingly, the maturation of Neisseria-containing phagosomes is also disturbed and cells lacking both LAMP proteins fail to kill the engulfed pathogens [159]. The maturation block caused by LAMP deficiency is at least partially due to the inability of autophagosomes and phagosomes to undergo dynein/dynactin-mediated centripetal movement along microtubules towards lysosomes [158]. Interestingly LAMP-2 single knockout mice show an impaired phagosomal maturation in neutrophilic leucocytes. The impairment of this innate immune defense mechanism leads to periodontitis, which is one of the most widespread infectious diseases worldwide. The retarded clearance of bacterial pathogens is due to an inefficient fusion between lysosomes and phagosomes, leading to less efficient killing of the ingested pathogens [160], [161]. Neutrophils of the LAMP-2 knockout mice also contain an accumulation of autophagic vacuoles [85], [160], which is likely also due to impaired fusion of autophagosomes with lysosomes.
Taken together these observations indicate that fusion with lysosomes is required to successfully complete both autophagosome and phagosome maturation that is necessary for efficient degradation of the cargo. Further, the results show that the maturation of autophagosomes and phagosomes share common features, because both processes are impaired with similar tissue and cell specificity in LAMP-2 deficient mice and LAMP-double deficient cell lines.
7. Conclusions
Degradation of cytosolic proteins in lysosomes via autophagy has turned out to have numerous, partly unexpected, roles in health and disease. Autophagy has been shown to contribute to innate and adaptive immunity and longevity, and to the prevention of cancer and neurodegeneration, just to mention a few of its newly-revealed functions. Treatments for human diseases that specifically target autophagy do not yet exist. It is likely, however, that such treatments will emerge in the future, once the molecular mechanisms of the processes involved in autophagy regulation have been clarified and suitable inducers and inhibitors for clinical trials have been identified.