These are steps, which if taken, may help recovery of the bodies abilities to utilise it’s elegant recovery mechanisms, to an elite level, relative to that which constrains us (age, history…).
1. First, the cornerstone for me:
Studies have shown, that Omega3 in various forms (Here is just a few days of searching the published medical papers for Flax), has a juggernaut of healing potential based in and around it.
My day, usually starts with 125g of Kefir Quark (called Kefir Protein in both Waitrose and M&S), blended with 62.5g of super fresh (very important as we’re dealing with Oxidation here) Flaxoil.
Above Picture, is when the Flaxoil is high lignan, if not the high lignan oil; then to be an official Budwig Breakfast, it should have freshly blended flax seeds.
This is blended with a hand blender, or whisk. Then I eat one spoonful alone, before having the rest with low glycemic fruit (Strawberries are my youngest daughters choice, Raspberries my eldest). Sometimes (almost always), I also have with this Planet Organic Paleo Vegan Granola:
Though this is not strictly Budwig kosher, if issues with Cancer or major illness I would not have this element personally.
This above step 1 is my absolute cornerstone with regards to fat metabolism, and protein intake, the sulfur rich proteins in the Kefir Quark has a natural affinity with the Flaxoil, which when blended make them water soluble, emulsify it to give a huge surface area, relative to having it just as an oil, a gestalt food where the sum exceeds the constituents hugely.
This is the foundation of Johanna Budwig’s research (the blend, forget the Granola), which she reported is the best was to re-ignite the Cell walls ability to take and carry Oxygen = Prevention of (or recovery from) Respiratory Acidosis.
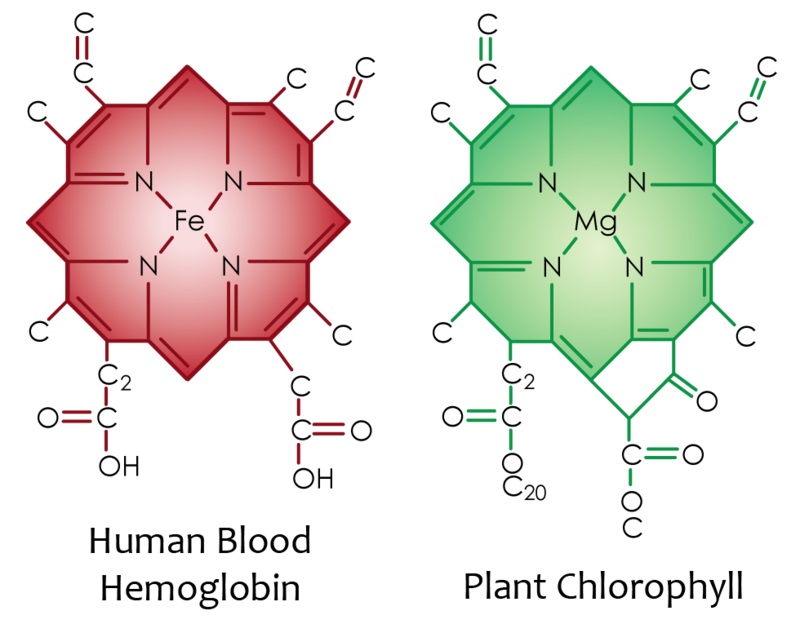
2. Green Smoothies – This is a two form deal. Firstly, to explain; the greens in leaves n vegetables etc. Are very rich in Chlorophylls, the centre of which is Magnesium:
So, as we can see, as well as the Magnesium boost you get, you also get a pre-constructed shell your body can use to apply Iron and you have Hemoglobin, then you have the folates, the VitaminK (which microbes in your body may convert to K2 if you eat correctly) and on and on.
When studying diet types, it became clear that green smoothies, with some oil as much of the goodness is fat soluble, creates a sensational dynamic for our Liver/Pancreas/Digestion/Well-Being.
How many of us can truly say we eat enough greens?
So, at least once a day, lately, I am trying to have the following:
1/3-1/4 Bag of Organic mixed leaves (M&S is my choice here) 1/3 Cucumber 1/3 Avocado 1 Tsp of Norwegian Kelp (Ascophyllum nodosum) 1 TBSP Flaxoil or Cold Pressed Organic Olive Oil 1 Small pinch of Epsom Salts 1 Tsp of Limewater or Calcium powder (I favour Min-Col, or, Cal II) 1 Handful of low Glycemic Fruit (Cherries today, very nice) 1 1/4 Lemon juice
Fill almost to top with clean water (I’ve got a distiller, but otherwise I like Evian, or Buxton is low Nitrate, Volvic is Silica rich and good if you have memory concerns), blend, and drink.
If no Lemon to hand, Apple Cider Vinegar, Raw with Mother, is an OK substitute, but Lemon is best IMO.
(Reams said the leaves and Olive Oil creates a gelatinous like substance to help lubricate our bowels, he advocated only a salad with olive oil for dinner, low late day protein to rest the body/liver/digestion and set yourself up for the following day, but this is a tough lifestyle adjustment to make, but dovetails research showing the autonomic nervous system needs to direct it’s juice to the Adrenals, key time 11pm-1am I believe, so it must be best to finish digesting proteins which take considerable oxygen, so it may direct it at your innate rest and repair cycles, such as recovering the adrenals, crucial in todays high processed, high refined, world)
3. Moreless Alkalising Drink:
1TBSP Limewater (This is Pickling Lime, but not sold in UK so make my own*) 1 TSP of Raw Honey (Organic, away from high pesticide areas), otherwise Unsulphured Molasses 3 TBSP Norwegian Kelp Powder 1 TBSP Apple Pectin 1/2 Cup Water 1/2 Juice Fresh Lemon 1 Pinch Epsom Salts
* This is made from Calcium Hydroxide, please understand not the powder itself, learn the method carefully, if not I have used Min-Col, Call II from Daily Manufacturing which are based on Reams teachings. Be Careful with Calcium!
Mix well, usually I mix this, except the lemon, leave it for an hour or two for the kelp and honey/molasses to fully mix easily, then juice in the lemon when I am to start sipping it. This is a high alkaline, and it is designed to be sipped upon hourly, to prop up your body (it’s well shown that most yeasts/viruses/bacteria/fungus of harmful nature are attracted to, and thrive, in more acidic terrain, mine runs acidic, my daughters when tested does, so this scales up as I see any of the multitude of signs for heightened acidity, in myself or my Girls, though my youngest rejects the Kelp as Yucky, but is fine with it without).
This above Alkaline mix, is the foundation of work, based upon studies of Biological Ionisation, which averts or prevents bodies which flirt with Metabolic Acidosis, Moreless would repeatedly say listen to your body and adjust up/down each ingredient, he rightly for his biochemistry expounded the virtues of all the trace minerals in Molasses, but I cannot tolerate that energetically so I use high, high quality raw organic honey, just enough to carry the Calciums and give a live Carbon bond to the otherwise slightly toxic hydroxide.
These are my foundations of well-being, on top of this, I try to eat as much fresh vegetables as I can. If we get to the stage that we are eating 50% raw fruit and vegetables (mainly veg, and low glycemic fruits), then we are giving such a rest to our Liver.
The above is a large part of the cornerstone of a diet, which if applied correctly, may help almost any illness, it is the foundational elements that the degradation of soil vitality and increasingly denatured living deprive us.
Understand what health is and what it means. Understand levels of body chemistry using urine and saliva. Understand the proper relationship of mental and spiritual aspects of health relating to body chemistry.
Throughout his research, Dr. Reams discovered that only two bodily fluids were needed to show your body chemistry levels – saliva and urine. If a person keeps his numbers in the Perfect Health or Healing range – it is believed the human body will maintain health. If the human body in not kept in the Perfect Health or in the Healing range – it is believed the body becomes diseased. The great news in pH testing is that you can manipulate your balances by controlling what you put into your body.
REAMS testing, this is how it works.
¬ The Carbohydrate measurement is made with a refractometer and it measures the number of brix in a urine specimen. It also represents the amount of potential energy available per pound of body weight. The ideal carbohydrate measure is 1.5 brix. Healing range is 1.2-2.0 brix. Below 1.2 represents low blood sugar. 5.5 and above represents borderline diabetes.
¬ The pH is a measurement of resistance and indicates the speed at which energy is moving through the body. A reading of 6.4 is the ideal speed for energy to move through the body. The pH is written as a fraction. The top number is the urine pH and the bottom number is the saliva pH. Healing range is 6.2-6.6. If you add the urine pH number to two times the saliva pH number and divide by three – the results will yield your average bodily pH. This is helpful in analyzing the direction of the overall pH of your body. Urine pH provides information about the blood, saliva pH provides information about the liver.
¬ The Salt or conductivity reading is ideally 6-7C. The conductivity number indicates the level of salts in the body. The salt number indicates whether the body has the correct number of electrolytes. Electrolyte levels indicate whether the body is undercharging or overcharging.
¬ Cell debris is an indication of the number of dead cells leaving the body. A sick body needs to rid itself of excess dead cells. The ideal cell debris number is .04M. The cell debris number tells how well the body is cooperating in the healing process. It is also the last number to come into balance.
¬ Urea readings are the Ammonia Nitrates and the Nitrate Nitrates added together. They equal the total ureas. Total ureas represent the total amount of unutilized protein that is being handled by the liver and sent to the kidneys for elimination.
¬ Cell Exchange Rate – Ultimately the entire body chemistry depends upon the correct cell exchange rate. Homeostasis – a new healthy cell is produced for each old cell dying off. If any of the numbers are off, the cell exchange rate is off and cells are not getting produced to maintain homeostasis. There are three classes of cells: Alpha Cells – perfect whole cells, Delta Cells – damaged or dead cells ready to be replaced, and Omega Cells – dead cells clumping and sticking together. Good health demands an even exchange rate – dead cells out and new cells in. If any of the numbers in the equation are out of range then the delta cells are not leaving the body. Any time the cell exchange is off, there is a mineral problem in the body. The body is made of minerals…The dust of the earth…Minerals are the basis for good health…You cannot build healthy cells without minerals.
¬ Mineral Assimilation is determined by the pH of the digestive system. A second issue is the atomic number of each of the minerals. The higher the frequency, the more difficult it is to assimilate the particular mineral.
What does this test reveal? Determines calcium needs for your body chemistry Tells what you are digesting or not digesting Tells if your body is assimilating nutrients Shows vitamin and mineral deficiencies Reveals if blood sugar is high, low or normal Will show if your body is supporting excess yeast candidiasis) or parasites Indicates if there is excess stress on internal organs such as the kidneys, liver, heart, colon or gall bladder Gives the health level of the liver and gall bladder Reveals if your body’s environment may be supporting: circulatory problems high blood pressure low blood pressure arthritis weight gain high cholesterol kidney/gall stones
PERFECT NUMBERS Through extensive research, Dr. Carey Reams discovered the “perfect numbers” for Biological Ionization, which represent the ideal cellular resistance required for life, just as 98.6 degrees represents the perfect resistance (temperature) for a healthy body. The higher the resistance, the higher the temperature. The lower the resistance, the colder the temperature. Death may occur from either extreme.
ENERGY FROM FOOD We do not live off the food we eat but off the energy produced by the food we eat. It is believed that we receive approximately 20% of our mineral energy from this digestive principal. Approximately 80% comes from the atmosphere. The more efficient the digestion, the more efficient the body is in extracting mineral energy from the air.
PREMATURE AGING Resistance is required for life. An excess of resistance can result in disease and death. Likewise, a deficiency of resistance can also result in disease and death. Dr. Reams determined that each of the 7 parameters, when all occurring simultaneously, represents the perfect 100% Metabolism Efficiency (the conversion of food into energy). The theory being that, if one could maintain a lifestyle that continually manifested the “perfect numbers”, there would be no premature aging.
BIOLOGICAL IONIZATION Of course there is no such perfect world, hence aging does occur. The objective, using “Reams Testing” as a guide, is to determine the proper lifestyle that allows a person to age, but not prematurely age. As the metabolism efficiency decreases, premature aging is more likely to occur, predisposing one to the disease process. How are urine and saliva tests different than a blood test? According to Dr. Reams, the blood changes every 15 minutes. The urine and saliva test was found to be more accurate. The testing of these two substances is amazingly accurate in determining the degree of wellness one might be experiencing and importantly what minerals, vitamins, and foods one should or should not eat. Biological Ionization Analysis is an excellent metabolic biofeedback device to indicate whether a particular lifestyle is beneficial or detrimental for any individual. It gives specific information on what vitamins and minerals are not being assimilated into the body’s cellular structure. It provides biofeedback on a holistic (emotional, physical, spiritual) level.
By Donald Kraus (Reams Practitioner) : http://www.bodylifedirect.com/
Lothar Hirneise: What is your fundamental research?
Dr Johanna Budwig: In 1950, I developed Paper Chromatography of fats. With this technique for first time fats, fatty acids and lipoproteins could be detected directly even in 0.1 ml of blood. Using Co 60 I was successful in producing the first differential reaction for fatty acids, and via radioiodine producing the first direct iodine value. I also developed control of atmosphere in closed system by using gas systems which act as antioxidants. Coloring, separating effects of fats and fatty acids were further developed. Behavior was studied in blue light, red light with fluorescent dyes.
I studied the electrical behavior of the unsaturated fatty acids with their “halo” using dyes with rhodamine red. With this technique I proved that electron rich highly unsaturated Linoleic and Linolenic fatty acids (found abundantly in flax oil) are the undiscovered decisive fats in respiratory enzyme function that Otto Warburg could not find. I studied the electromotoric function of pi-electrons of the linolenic acid in the cell membranes, for all nerve function, secretions, mitosis, as well as cell breakdown. I also examined the synergism of the sulfur containing protein with the Pi-electrons of the highly unsaturated fatty acids and their significance for the formation of the hydrogen bridge between fat and protein, which represent “the only path” for fast and focused transport of electrons during respiration.
This immediately caused a furor. Cancer problem was brought in. Hydrogenated fats, which includes all Trans fatty acids proved to be respiratory poisons. This was extensively published in 1950 in many journals including “New Directions in Fat Research.”
Lothar Hirneise: What is the prime cause of Cancer?
Dr Johanna Budwig: In 1928 Dr. Otto Warburg proved that all normal cells have an absolute requirement for oxygen, but cancer cells can live without oxygen – a rule without exception. Deprive a cell 35% of its oxygen for 48 hours and it may become cancerous. Dr. Otto Warburg has made it clear that the root cause of cancer is oxygen deficiency in the cells, which creates an acidic state in the human body.
He also discovered that cancer cells are anaerobic (do not breathe oxygen), get the energy by fermenting glucose producing lactic acid and cannot survive in the presence of high levels of oxygen. Long back in 1911 Swedish scientist Torsten Thunberg postulated that sulfur containing protein (found in cottage cheese) and some unknown fat is required to attract oxygen in the cell. This fat plays a major role in the cellular respiration. For nearly half century scientists were trying to identify this unknown and mysterious fat but nobody succeeded.
Lothar Hirneise: How did you develop cancer therapy which is called Budwig Protocol?
Dr Johanna Budwig: During my research I found that the blood of seriously ill cancer patients was deficient in these unsaturated omega-3 fats (Linoleic and Linolenic fatty acids), lipoproteins, phosphatides, and hemoglobin. In addition, I noticed that cancer patients had a strange greenish-yellow substance in their blood that I could not find in the blood of healthy people. Then I wanted to develop healing program for cancer.
So I decided to straight waygo for human trials and started to give flax oil and cottage cheese to the cancer patients from four big hospitals in Münster. After just three months, patients began to improve in health and strength, the yellow green substance in their blood began to disappear, tumors gradually receded and at the same time as the nutrients began to rise. Thus I had a cure for cancer. It was a great victory and the first milestone in the battle against cancer. My treatment is based on the consumption of flax seed oil with low fat cottage cheese, raw organic diet, detoxification, mild exercise, flax oil massage and the healing powers of the sun. I have treated approx. 2500 cancer patients during last few decades. Prof. Halme of surgery clinic in Helsinki used to keep records of my patients. According to him my success was over 90% and this was achieved in cases where allopathy failed.
Lothar Hirneise: Can you tell us more about the unsaturated fatty acids and their net-like connections?
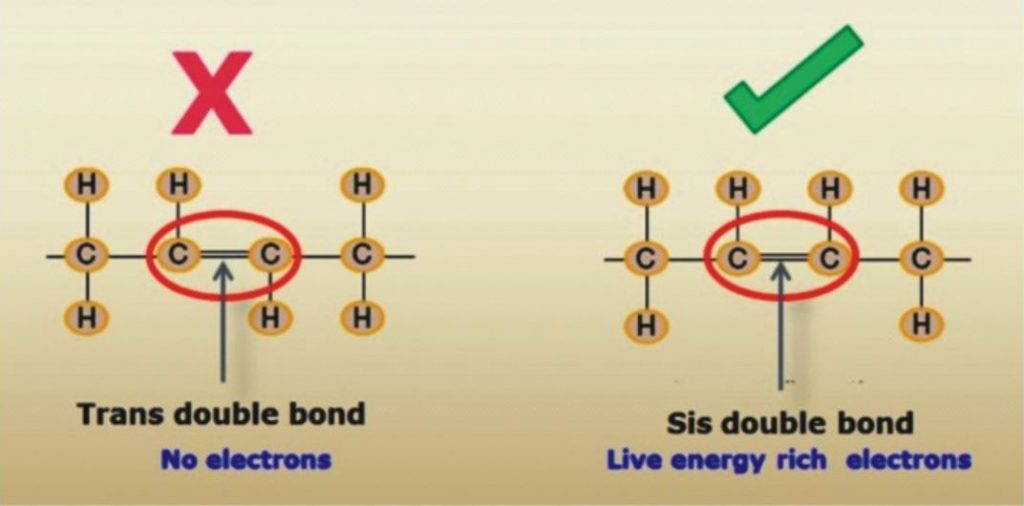
Dr. Johanna Budwig: Fatty acid is a carboxylic acid having unbranched chain of 4 to 28 carbons. The saturated fatty acids have primarily short carbon chains. In butter, coconut fat, goat fat and sheep fat the fatty acids consists of 4, 6, 8, 10 or 12 carbons, are saturated, however they are also easily co-combustible if the essential fatty acids are present. The unsaturated vital fatty acids really start with the chain with 18 carbon compounds. There are also fatty acids with up to 30 links. Fatty acids with 18 links, like in flax oil with the higher level of unsaturation, are more important for human, particularly for the brain functions of man. Linoleic acid rich in electrons is considered vital. There is a particularly high amount of energy in this double double bond of the linoleic acid. This energy wanders and is not fixed in place as it is with a chemical compound, such as with table salt. This energy, wandering between electrons and the positively charged protein with sulfur groups is an alternating association process in the electromagnetic field. This is very important. Perhaps you are familiar with the painting of Michelangelo, where God creates Adam (two fingers pointing to each other, however they do not touch). This is quantum physics, the fingers do not touch. The physicists who I know, Max Planck, or Albert Einstein, or Dessauer all represent the view that man is created by God in His image. You see in being together as human beings there is certainly also a connection without directly touching the other person. The dipolarity with a single double bond in olive oil is weaker than it is in sunflower seed oil, which is has two double bonds.
This double double bond is considered to be vital for man. However if the same chain length of 18 carbons has three unsaturated fatty acid compounds, then the electrical energy is as strong as a magnet. This electronic energy is negatively charged. The positively charged sulfur groups of the protein adhere in the unsaturated bonds where the electrons are and that is where they insert their sulfur-containing compounds.
This produces the lipoproteins. The life process is sustained in the interplay between the positively-charged particles and negatively-charged particles. In this process there is no connection, and this is our life element. If radical damage occurs at this point through fatty acids that has lost electron energy, but rather are cross-linked like a net, then the dipolarity can no longer work actively in this net. This is the deadly effect of radicals, because instead of the chains with the electron clouds they interlace a net without electron clouds, indeed with unsaturated bonds, but without dipolarity. I quickly knew that the triple unsaturated fatty acids, which were called linolenic acid, and which no one had isolated before me, had 18 links and that they did not always carry their double bonds at the same point. They have such a strong electronic energy compared to the heavier matter in the 18-link fatty acid chains, that biologically this energy is far greater than it is with the next arachidonic acid with 20 links. The highest electron collection is with the combination of linoleic-linolenic fatty acids in flax oil. The linolenic acid as conjugated (interaction of neighboring double bonds in the molecule that are separated by a single bond) fatty acid is even more effective and is even more strongly interplay with linoleic acid as it is present in the flax oil for oxygen absorption. This was relatively easy for me to verify in my experiments. I would like to emphasize this. The combination of double unsaturated linoleic acid with triple unsaturated linolenic acid is particularly well-combined in flaxseed.
Lothar Hirneise: Is it this energy that heals cancer?
Dr. Johanna Budwig: Yes, this energy is now movable and it is easily released. It is precisely this energy that heals cancer, or does not even allow it to occur. If this vital element is present then no tumor can exist. This vital element is a deciding factor in the immune system. There is no effective factor in the immune system other than the essential fatty acids.
Lothar Hirneise: What is an electron cloud?
Dr Johanna Budwig: If the enhancement of electronic energy is always higher through absorption of sun photons in the unsaturated fatty acids e.g. in linolenic fatty acids, then the power of the electrons is so high in the dipolarity in between gravity and electrons, that they lifts off of the heavy mass and floats like a cloud hence I called them electron cloud.
Lothar Hirneise: What is the significance of the cloud?
Dr. Johanna Budwig: No life form has as much energy to store the electrons and photons as does man. The electronic energy stored particularly in the vital, highly unsaturated fatty acids, is very strong life element for man. Man cannot live without them. If oils are treated with heat and harsh chemicals (during refining and hydrogenation process to increase their shelf life) then the wealth of vital electronic energy is destroyed and Trans fats are formed with net like connections. They are no longer vital fats with 18 links, but rather they form cross-links between the fatty acids like a large net, and are highly damaging to our body, do not adhare with proteins, do not attract oxygen and act like a radicals. I repeat because it is so important: I have detected particles in oils treated with steam, which indeed have a positive iodine value, but which are highly toxic for man.
Lothar Hirneise: So you preach against these toxic hydrogenated and refined oils?
Dr. Johanna Budwig: I am completely against using these “pseudo” fats – “hydrogenated”, “partially hydrogenated”. These are the biggest enemy of mankind. I had scientific proofs. The heart rejects these fats and they are deposited as inorganic fat on the heart muscle itself. They end up blocking circulation, damage heart action, inhibit cell renewal and impede the free flow of blood and lymph fluids.
But it was highly profitable business for multinationals. When I preached against these fats, they stood against me, first they tried to give me hush money and when I refused they filed many fake court cases against me. I was working for humanity and had scientific proof.I was like rock of Gibraltar in my decision; I fought and won all the cases ultimately.
Lothar Hirneise: What is mysterious marriage of sun and moon in terms of quantum physics?
Dr. Johanna Budwig: The photon, the quantum, is the smallest component of the sunlight. It is the fastest moving traveler, it speeds along with time. The photon is always in motion. Nothing can ever halt its motion. The photon is full of all colors, and can change its color and its frequency. The photon is the purest form of energy, the purest wave, can unite with a second photon when it is in resonance with the other, to form a “short-lived particle.” This particle, known as a p0 particle, can break up into two photons again, without mass, as a pure wave in motion. This is the basis for the wonderful back and forth movement between light and matter. The photon can never be pinned down to one location. This is the foundation for the Theory of Relativity.
This very active, dynamic and energetic photon can be captured by electron that are in resonance with the photon. What does that mean? Electrons are already a component of matter. They are also continuously in motion. They constantly oscillate on their own wavelength. They have their own frequency, just like a radio receiver that is set to a specific wavelength.
The electron loves photons. It attracts photons by its magnetic field. When an electrical charge moves, it always produces a magnetic field. The moving photon also has a magnetic field. Both fields attract each other when the wavelengths are in tune. The wave length of the photon – which the photon can change – must fit into the wavelength of the orbiting electron so that the orbit maintains a complete wavelength. This is eternal love between sun-god, photons and moon-goddess, electrons, this is the mysterious marriage of the sun and moon in terms of modern science.This feature is extremely interesting in terms of its physical manifestation, its biological and even its philosophical consequences. All matter has its own inherent vibration. Of course, this also applies to living substances. The energy which is being absorbed must correspond to the inherent wavelength.
The sun rays are very much in harmony with humans. It is no coincidence that we love the sun. The resonance in our biological tissue is so strongly tuned to the absorption of solar energy that there is nothing on earth that has a higher concentration of solar energy photons than humans. This enrichment with solar energy depends strongly on the “like energy” aspects, a wavelength that is compatible with humans, and this is supported when we eat foods that have electrons with electromagnetic fields that attract the photons of the sun. An abundance of these electrons, which are tuned to the solar energy frequency, exist, in many seed-oils. Scientifically these oils have even been designated as electron-rich, “essential,” highly unsaturated fats. But when we began processing fats to increase the shelf life, nobody thought about the significance that this would have for the survival and the further development of the human species. We destroyed their extremely important wealth of electrons, which are very mobile and react so wonderfully to sunlight.
Lothar Hirneise: What is the concept of “human” and “anti-human” in terms of quantum physics?
Dr. Johanna Budwig: It is interesting that in the science of quantum physics the concept of “human” has already been coined. It is “human”, with the highest accumulation of photons, always striving toward the future, who possesses within himself the highest potency of solar energy on the earth. The physicists project from mathematical equations that man, with his wealth of electrons, is directed forwards future in time.
Using mathematical equations that are valid in physics, reversing the time quotient represents the mirror image of man — the “anti-human,” whereas “man” represents the picture of highest rank in terms of physics, i.e. directed against entropy. The “anti-human” is directed back in time. The “anti-human” possesses few solar energy photons, in physical terms electron-poor, directed into the past — also in his thinking — is paralyzed in his life functions, lacks energy and strength because he is missing the electrons that are in harmony with the sun as “life-element.”
The physical processes which are generated through the use of X-rays, gamma rays, atom bombs, or cobalt radiation, are pointed in the same direction as the development toward the “anti-human,” from the perspective of physics and mathematics. The electron structure of the life functions is destroyed by these rays. According to the so-called “World Line” and the Theory of Relativity of modern physics, time and space are connected together in one equation. It is very interesting to investigate our food from this perspective. Fats that have had their electron structure destroyed to make them keep longerpromote the development of the “anti-human”and have a very detrimental effect on the future-directed, electron-rich human being, according to the “World Line diagram.” They disturb the electron exchange within living tissue because they, like tar, act as insulators against electrical conductivity, plainly deaden the life functions at the respective operative locations, e.g. in organs, and in growth centers of the body, as well as throughout the whole body. They also promotes cancer.
On the other hand, electron-rich nutrition, electron-rich highly unsaturated oils,herbs and fruits which are rich in aromatics and natural color components that correspond to the colors of the photon s of sunlight — all these increase the absorption, storage and utilization of the sun’s energy.
Lothar Hirneise: What is your view point about surgery for tumors?
Dr. Johanna Budwig: I am totally against radiation and chemo; I also reject hormonal treatment. Surgery must be considered individually. I am not a proponent of quickly making artificial anus. Conventional oncology no longer does justice to the cancer patients.
Lothar Hirneise: You also studied medicine at the age of 47 years.
Dr. Johanna Budwig: (smiling) Yes handsome! That’s right, my opponents were accusing me that how can I treat cancer patient without doctors degree. This thing pinched me, so in 1955,I joined medical school in Göttingen. There I was using my therapy very successfully in various clinics. I still remember the time I was working late one night in Göttingen, a woman came to me, with her small child whose arm was supposed to be amputated due to a tumor. I treated her and soon the subject of amputation was dismissed and the child quickly did very well.
Because I was still a medical student at this time, I was summoned to appear before the Municipal Court due to a petition that I should be prohibited from studying medicine. I explained the truth in the court. The judge rejected the case and said, “You have done a good job. In my area of jurisdiction nothing will happen to you. If it does there will be a scandal in the scientificcommunity.”
Lothar Hirneise: What do you recommend for prevention of cancer?
Dr. Johanna Budwig: Only flax oil as oil. I reject frozen and preserved meat. Fresh meat is OK. No frozen food and no bakery products. Oleolux should be used as butter. Prepare fruit juices yourself. Cheese and potatoes are OK. Also the electromagnetic environment (e.g. microwave and mobile phones etc.) in which we live is very important. I reject synthetic textiles and foam mattresses because they steal lot of forces from you. A lot of wood in home construction and woolen or silk carpets are also important. Wear gemstones, they also have good biological radiation. The environment and living conditions must be as biological (organic & natural) as possible. Regular sleep is very important.
Here are some excerpts of a paper on the functioning of Hydroxychloroquine, the drug being touted as a medicine to combat CoronaVirus. It can be seen that it’s major action, supportive of the previous data I’ve presented, is raising pH…
Hydroxychloroquine (HCQ) enters and accumulates in lysosomes along a pH gradient. In lysosomes, hydroxychloroquine inhibits the degradation of cargo derived externally (via endocytosis or phagocytosis) or internally (via the autophagy pathway) in autolysosomes by increasing the pH to prevent the activity of lysosomal enzymes. Inhibition of lysosomal activity can prevent MHC class II-mediated autoantigen presentation.
Hydroxychloroquine can also accumulate in endosomes and bind to the minor groove of double-stranded DNA. This drug can inhibit Toll-like receptor (TLR) signalling by altering the pH of endosomes (involved in TLR processing) and/or preventing TLR7 and TLR9 from binding their ligands (RNA and DNA, respectively). Hydroxychloroquine can also inhibit the activity of the nucleic acid sensor cyclic GMP-AMP (cGAMP) synthase (cGAS) by interfering with its binding to cytosolic DNA. By preventing TLR signalling and cGAS–stimulator of interferon genes (STING) signalling, hydroxychloroquine can reduce the production of pro-inflammatory cytokines, including type I interferons.
….
As the pH in lysosomes is optimal for lysosomal enzymes involved in hydrolysis, by increasing the pH of endosomal compartments85, chloroquine and hydroxychloroquine might impair the maturation of lysosomes and autophagosomes and inhibit antigen presentation along the lysosomal pathway (Fig. 3).
…
These processes possibly occur, at least in part, through drug-mediated changes in the pH of autophagosomes and/or lysosomes, which indirectly influence immune activation; however, such a mode of action requires additional validation to aid with future drug development.
This is the drug they are saying shows great promise for Coronavirus treatment.
Here is excerpts from a paper just being released, where studies have shown 100% cure rate with CoronaVirus:
He talks about lowering the acidity in the cells…
https://www.covidtrial.io/
From their prelim paper:
Specifically, the CDC research was completed in primate cells using chloroquine’s well known function of elevating endosomal pH. The results show that “We have identified chloroquine as an effective antiviral agent for SARS-CoV in cell culture conditions, as evidenced by its inhibitory effect when the drug was added prior to infection or after the initiation and establishment of infection. The fact that chloroquine exerts an antiviral effect during pre- and postinfection conditions suggest that it is likely to have both prophylactic and therapeutic advantages.
…
When chloroquine is added after infection, it can rapidly raise the pH and subvert on-going fusion events between virus and endosomes, thus inhibiting the infection. When added after the initiation of infection, it likely affects the endosome-mediated fusion, subsequent virus replication, or assembly and release. Specifically, rapid elevation of endosomal pH and abrogation of virus-endosome fusion may be the primary mechanism by which virus infection is prevented under post-treatment conditions.
…
Specifically, the virus depends on turning over the host proteins to trigger response for available building blocks to make their own proteins or nucleic acids. They break down due to low PH catalyzed by hydrolysis.
…
It is this part of the coronavirus’ replicative path that chloroquine inhibits. Notably, because of its nitrogen structure, chloroquine has the unique ability to get into cells and cross endosomal membranes. Once inside, nitrogens in chloroquine (and quinines in general) prevent acidification by absorbing a high amount of hydrogens that simply then interact with nitrogen and then chloroquine becomes positively charged – an ionic interaction which makes it harder for the endosome to become acidified. The result is a buffer that holds it at the higher pH and prevents it from becoming acidic enough to be functional. To summarize, because chloroquine has a multitude of extra nitrogens, once it crosses the membrane and enters an organelle, the organelle is prevented from reaching a lower pH. The organelle’s enzymes cannot work because the donor group will be a hydrogen ion, disabling the hydrolysis required for coronavirus replication. This means that all kinds of events in the cell are incapable of performing optimally, including viral replication.
Bill brought inhalation of H2O2 to the publics attention, he used it 5-6 pumps, 6 or 7x a day. To keep his Oxygen levels high, to keep the Viruses out, which come when Oxygen is down.
This was something I tried many years ago, but didn’t feel a direct or urgent need to do, so put it into my memory locker.
However, after studying this new Viral Epidemic which increasingly surrounds us, I pulled the information back to my short term memory to mix with the pH principles I had been reflecting on.
My daughter has suffered from Chronic Lung Disease as a result of being born at 24weeks gestation, so Oxygen and Viral susceptibility has been something I am used to reflecting deeply upon.
Anyway, the data shows that if you are of a lower pH, this virus group (Corona) is 10x more forceful and you are 10x more susceptible (at a pH of 6 Vs 7), so after ensuring I got my diet plans and supplements into order, I began to reflect over and over how else I can best prepare.
This is when Bill Munro came back to my mind, so I ordered 2x Nasal Vaporiser’s for about £3, and I decided that I would start taking a similar number of rounds of H2O2 as Bill did, but diluted to 1.5% with 50% Distilled water and 50% Food Grade H2O3 at 3%.
The literature for killing the Virus shows that 0.5% H2O2 kills the virus in 1 Minute, so 1.5% should be effective in 20 Seconds or thereabouts. So, to my unqualified mind, this leads me to presume that several rounds of this daily may safeguard me against this threat, if not preventing it from entering my airways entirely, then at least killing it off 5-6 times a day, minimising the intensity of the infection as best I may.
When I have talked of this method to others, peoples minds immediately seem to jump to “you are swallowing bleach”, with a shocked reaction, yet I explain that this is simply water with an extra Oxygen molecule attached, which people regularly treat their teeth with at 7% some 450% stronger than I am using. People seem to not bat an eyelid at using 4.5x stronger bleach for their teeth and vanity, yet to stave of seemingly very harsh virus people react like I am the crazy one!
This method along with Alkalising my terrain via diet and supplements are the best methods I have encountered to protect myself against this horrid viral outbreak.
If the outbreak worsens in the UK, I am intending to upgrade to the full 3%
Authors : Li Y, Wang J , Yue J , Wang Y, Yang C, Cui Q
Cell Biology International : 2018 February
Abstract
Magnesium, as a physiological calcium antagonist, plays a vital role in the bone metabolism and the balance between magnesium and calcium is crucial in bone physiology. We recently demonstrated that matrix mineralization in human bone marrow-derived mesenchymal stem cells (hBMSCs) can be suppressed by high Mg2+ .
However, a complete understanding of the mechanisms involved still remains to be elucidated. As mitochondrial calcium phosphate granules depletion manifests concurrently with the appearance of matrix vesicles (MVs) and autophagy are associated with matrix mineralization, we studied the effect of high extracellular Mg2+ on these pathways.
Our results first demonstrated that high Mg2+ has a significant inhibitory effect on the generalization of extracellular mineral aggregates and the expression of collagen 1 along which the mineral crystals grow. Transmission electron microscope results showed that less amount of MVs were observed inside hBMSCs treated with high Mg2+ and high Mg2+ inhibited the release of MVs.
In addition, high Mg2+ significantly suppressed mitochondrial Ca2+ accumulation. Autophagy is promoted as a response to osteogenesis of hBMSCs. High Mg2+ inhibited the level of autophagy upon osteogenesis and autophagy inhibitor 3-MA significantly suppressed mineralization. Exogenous ATP can reverse the inhibitory effect of high Mg2+ by increasing the level of autophagy.
Taken together, our results indicate that high Mg2+ may modulate MVs-mediated mineralization via suppressing mitochondrial Ca2+ intensity and regulates autophagy of hBMSCs upon osteogenesis, resulting in decreased extracellular mineralized matrix deposition. Our results contribute to the understanding of the role of magnesium homeostasis in osteoporosis and the design of magnesium alloys.
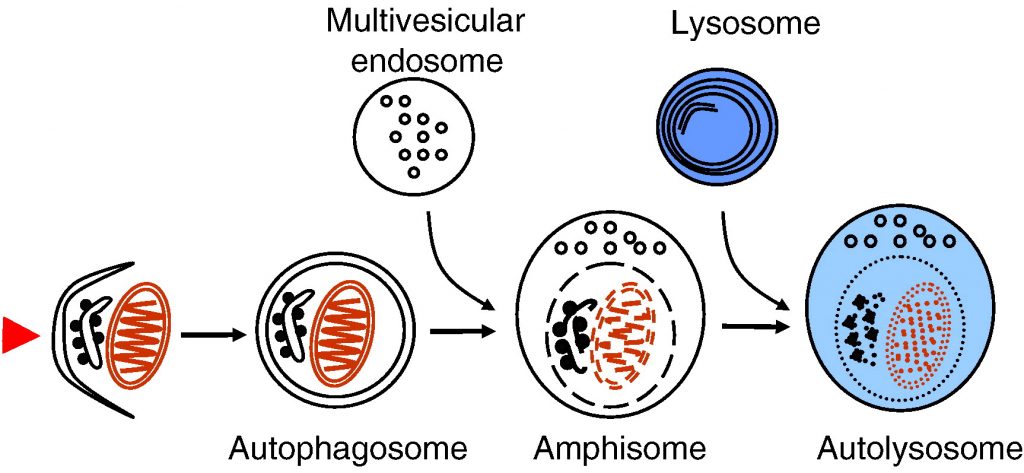
Autophagy delivers cytoplasmic material and organelles to lysosomes for degradation. The formation of autophagosomes is controlled by a specific set of autophagy genes called atg genes. The magnitude of autophagosome formation is tightly regulated by intracellular and extracellular amino acid concentrations and ATP levels via signaling pathways that include the nutrient sensing kinase TOR. Autophagy functions as a stress response that is upregulated by starvation, oxidative stress, or other harmful conditions. Remarkably, autophagy has been shown to possess important housekeeping and quality control functions that contribute to health and longevity. Autophagy plays a role in innate and adaptive immunity, programmed cell death, as well as prevention of cancer, neurodegeneration and aging. In addition, impaired autophagic degradation contributes to the pathogenesis of several human diseases including lysosomal storage disorders and muscle diseases.
Fig. 1. Schematic presentation of autophagosome formation and maturation by fusion with endosomes and lysosomes. The arrowhead on the left illustrates the induction signal that initiates the process.
5. Physiological functions of autophagy
5.1. Stress response and housekeeping function
The role of autophagy as a survival mechanism during short-term amino acid starvation is well documented. Macroautophagy is induced by starvation of serum and amino acids; autophagosomes can be detected already after 15–30 min of starvation [55]. During long-term starvation, chaperone-mediated autophagy increases and macroautophagy decreases [80], [81]. Yeast strains defective in autophagy do not survive nitrogen starvation [38]. Knockout mice deficient in one of the central autophagy proteins, Atg5, show that autophagy is indispensable for the energy metabolism immediately after birth [82]. Atg5 knockout mice die of starvation one day after birth.
In muscle and heart cells, autophagy seems to have a special housekeeping role in the turnover of cytoplasmic constituents including mitochondria. This is revealed by myopathy and cardiomyopathy in patients and mice possessing a defective autophagic degradation due to deficiency of the lysosomal membrane protein LAMP-2 [83], [84], [85]. LAMP-2 deficiency is described in detail later in this review. The importance of autophagy for the heart muscle is supported by a study showing that heart-specific loss of the autophagy protein Atg5 causes cardiomyopathy in mice [86]. Evidence has been published suggesting that damaged mitochondria might be autophagocytosed selectively in a process termed mitophagy [87]. Mitochondria are the major source of reactive oxygen species in cells. Interestingly, reactive oxygen species are necessary for the signal transduction pathway initiating starvation-induced autophagy [88].
It was proposed long ago that autophagy has a role in growth regulation, as suggested by decreased autophagy during growth of the kidney after unilateral nephrectomy [89]. Inducible knockdown of the autophagy protein Atg5 in cell culture shows that autophagy negatively controls cell size [90]. Similar result was observed in Drosophila fat body over-expressing the autophagy protein Atg1 [91].
Autophagy contributes to intracellular quality control and housekeeping, especially in turnover of aggregate-prone proteins. Prevention of autophagy by conditional knockout of atg7 leads to accumulation of ubiquitinated protein aggregates in mouse tissues [92]. Tissue-specific knockout of autophagy proteins in the central nervous system causes accumulation of ubiquitin-positive protein aggregates and neurodegeneration in mice [93], [94]. Further, enhanced autophagy reduces the toxicity of the Huntingtin protein aggregates that accumulate in Huntington disease [95]. Autophagy may prevent aggregate formation by degrading the proteins as monomers, oligomers, or after aggregate formation [96]. It is not clear at present whether aggregated proteins are segregated preferentially, or whether they are removed via unspecific autophagic uptake of cytoplasm. Two proteins have been proposed to function during the uptake of protein aggregates: Alfy and p62 [97], [98]. p62 binds to both ubiquitin-conjugated aggregate-prone proteins and the autophagosome protein LC3 [99], which suggests that it could selectively recruit autophagy machinery to the aggregates and enhance their autophagic clearance.
In addition to removal of cytoplasmic aggregate-prone proteins, autophagy also contributes to the quality control in the ER. Unfolded protein response induces autophagy, and this induction is beneficial for cell survival [100], [101], [102].
Autophagic degradation is also needed for early embryonic development. A recent study shows that autophagy-defective mouse eggs fertilized with autophagy-defective sperm, failed to develop beyond the four and eight cell stages [103]. The authors suggest that autophagy may be needed in the preimplantation embryos for protein recycling, production of amino acids for protein synthesis or substrates for energy production, or for removal of obsolete maternal factors.
5.2. Innate and adaptive immunity
Autophagy contributes to both innate and adaptive immunity [104], [105]. In some cases autophagy can protect cells against intracellular pathogens. Induction of autophagy during Herpes simplex virus infection, and localization of viral particles inside autophagic vacuoles, were proposed to indicate that autophagy acts as a host-defense mechanism in infected cells [59]. The Herpes virus virulence protein, ICP34.5, inhibits autophagy, suggesting that the virus has developed a way to prevent the autophagic defense of the host cell. Autophagy may also help cells to defend against intracellular bacteria [106]. Sequestration of intracellular Group A Streptococci in autophagosome-like structures protects cells against the bacteria [107]. Mycobacterium tuberculosis is normally able to survive inside macrophages by preventing the fusion of phagosomes with lysosomes. Surprisingly, induction of autophagy bypasses the maturation defect, leading to phagolysosome formation and bacterial killing [108].
Macroautophagy also contributes to antigen presentation. Major histocompatibility complex (MHC) class II molecules present products of lysosomal proteolysis to CD4(+) T cells. Extracellular antigen uptake is considered to be the main source of MHC class II ligands. However, it was demonstrated that in MHC class II-positive cells, including dendritic cells, B cells, and epithelial cells, autophagosomes continuously fuse with multivesicular MHC class II-loading compartments [109]. This pathway is of functional relevance, because targeting of the influenza matrix protein 1 to autophagosomes enhances its MHC class II presentation to CD4(+) T cells. Thus it seems that macroautophagy efficiently delivers cytosolic proteins for MHC class II presentation and can improve helper T cell stimulation.
5.3. Cell death
Autophagy also seems to have roles in programmed cell death [110], [111]. Type II programmed cell death, or autophagic cell death, was originally described in mammary carcinoma cells [112], [113]. Autophagy proteins were shown to be necessary for cell death under certain conditions, such as in apoptosis-defective cells [114], [115], [116]. In this scenario autophagy is needed for the execution of cell death. Under other conditions, such as nutrient starvation, autophagy protects cells against apoptosis by providing nutrients [117], [118], [119]. The regulation of apoptosis and autophagy are linked via the antiapoptotic protein Bcl-2. Bcl-2 inhibits Beclin 1-dependent autophagy by binding to Beclin 1 and preventing its association with Vps34 [52]. This anti-autophagy function of Bcl-2 was proposed to help maintain autophagy at levels that are compatible with cell survival, rather than cell death.
Lipids may also regulate autophagy and its outcome to the host cell. Ceramide and sphingosine 1-phosphate, a metabolite of ceramide, both induce autophagy in mammalian cells [120]. The outcome on cell survival is however different: ceramide promotes cell death, while sphingosine 1-phosphate increases cell survival. Ceramide is part of the signaling cascade initiated by chemotherapy, while sphingosine 1-phosphate is part of the signaling cascade initiated by starvation. Beclin 1 level and the autophagy response are stronger during ceramide signaling [121].
Autophagy has functions in cell death during development [122]. atg genes are necessary for the clearance of apoptotic cells during embryonic development in mice [123]. Autophagy contributes to dead-cell clearance during programmed cell death by maintaining cellular energy levels in the dying cells, thereby allowing the generation of cell surface and secreted signals that then promote engulfment of cell corpses by neighboring cells. Autophagy is also indispensable for the execution of certain types of cell death during development. The degradation of Drosophila salivary glands by type II programmed cell death depends on autophagy [73].
5.4. Aging and longevity
Finally, autophagy also contributes to longevity [124]. Reduced caloric intake increases longevity in several animal species. Increased autophagic turnover of cytoplasmic constituents including mitochondria was shown to contribute to the longer life in the dieting animals [125]. Further evidence that autophagy contributes to longevity come from Caenorhabditis elegans mutants possessing a defective insulin receptor (daf2 mutant), which live longer than control worms. The increased lifetime of these mutant worms depends on a functional autophagic pathway [126]. Moreover, knockdown of autophagy gene products including Atg7 and Atg12 were shown to shorten the lifespan of both wild type and daf2 mutant C. elegans[127]. Further, promoting basal levels of autophagy in the nervous system of adult Drosophila enhances longevity of the flies [128]. Together these studies give strong support for a role of autophagy in the prevention of aging.
Fig. 6 summarizes the physiological functions of autophagy described above.
Fig. 6. A summary of the functions of autophagy in health and disease.
6. Autophagy and disease
6.1. Cancer
Impaired autophagy contributes to cancer development [2], [129], [130]. Beclin 1 is monoallelically deleted in a large proportion of human breast and ovarian cancers. Over-expression of Beclin 1 in a breast cancer cell line increases autophagy and decreases the growth and tumorigenicity of these cells [131]. Mice with heterozygous deletion of Beclin 1 have less autophagy and more tumors than control mice [132], [133]. Further, the other autophagy-promoting components of the Beclin 1/Vps34 complex, UVRAG and Ambra 1 (Fig. 4), are also tumor suppressors [48], [49]. Moreover, knockout of Bif-1, also part of the Beclin 1 complex, significantly enhances the development of spontaneous tumors in mice [50]. On the other hand, binding of the proto-oncogenic proteins Bcl-2 or Bcl-XL to Beclin 1 inhibit autophagy [51], [52]. In addition to the Beclin 1 complex, other tumor suppressors also enhance autophagy. PTEN is a phosphatase that decreases the concentration of class I PI3 kinase product and enhances autophagy [72]. PTEN is also a tumor suppressor [134]. Further, the activities of Ras and class I PI3-kinases inhibit autophagy and promote cell growth. Ras is mutated and class I PI3 kinases are upregulated in many cancers [135], [136].
The results described above show that autophagy contributes to the prevention of tumorigenesis. Impaired autophagy can contribute to tumor formation via impaired regulation of cell growth, and/or via decreased cell death. In addition, it was shown that failure to sustain metabolism via autophagy results in increased DNA damage. This chromosomal instability increases tumor progression [137].
In advanced cancers, autophagy may have the opposite effect on the tumor development. Autophagy can benefit the progression of the tumor because it can provide nutrients during starvation [129], [130], [138]. In addition, autophagy was recently shown to improve the survival of p53-deficient cancer cells under starvation or hypoxic conditions [139]. These findings suggest that autophagy inhibition, rather than stimulation, might be beneficial in treatment of advanced cancer.
6.2. Neurodegeneration
Many age-related neurodegenerative diseases are characterized by the accumulation of ubiquitin-positive protein aggregates in affected brain regions. These misfolded, aberrant proteins can disrupt neuronal function and cause neurodegeneration. As described earlier, autophagy is necessary for the clearance of aggregate-prone proteins that are toxic especially for post-mitotic cells like neurons [130]. Tissue-specific knockout of the autophagy genes in neurons causes a massive accumulation of ubiquitin-positive protein aggregates and neurodegeneration in mice [93], [94], indicating that autophagy is needed for the constitutive clearance of aggregate-prone proteins. Autophagy was recently shown to enhance the clearance of Huntingtin, mutant tau, synphilin 1 and α-synuclein, but not AIMP2 (p38) and mutant desmin [140]. This study indicates that autophagy is not able to degrade all protein aggregates. However, the role of autophagy has been demonstrated in Huntington’s disease, caused by mutations in Huntingtin, and familial Parkinson’s disease, caused by mutations in α-synuclein. Enhanced autophagy in animal models of these diseases improves clearance of the aggregated proteins and reduces the symptoms of neurodegeneration [95], [141].
ESCRT complexes are necessary for the biogenesis of multivesicular endosomes. As described earlier, multivesicular endosomes are necessary for the maturation of autophagosomes into degradative autolysosomes. Mutations in ESCRT III subunits CHMP2B or mSnf7-2 are associated with two neurodegenerative diseases, frontotemporal dementia and amyotropic lateral sclerosis. Both diseases are characterized by abnormal ubiquitin-positive protein deposits in affected neurons. Cell lines and fruit flies depleted of CHMP2B or mSnf7-2 show decreased autophagic degradation, increased levels of ubiquitin-positive aggregates and increased neurodegeneration [27], [29].
Alzheimer’s disease is characterized by the accumulation of extracellular amyloid plaques in the brain. These plaques consist of aggregated β-amyloid (Aβ) peptide. Autophagy was proposed to contribute to the production of Aβ. Autophagic compartments containing both amyloid precursor protein and Aβ accumulate in dystrophic neurons in Alzheimer brain [142], [143]. Purified autophagic vacuoles contain all necessary constituents for Aβ production [142], and autophagic compartments were identified as a major reservoir of intracellular Aβ in the brain of Alzheimer patients and mouse models. The primary cause for the increased accumulation of autophagic compartments in Alzheimer’s disease was recently suggested to be their retarded maturation to autolysosomes [144].
A recent study, however, challenges the idea that autophagy contributes to the pathogenesis of Alzheimer’s disease. Beclin 1 was shown to be decreased in affected brain regions of patients with Alzheimer disease early in the disease process [145]. Heterozygous deletion of Beclin 1 in mice decreased neuronal autophagy and resulted in neurodegeneration. Transgenic mice expressing human amyloid precursor protein have been used as a mouse model for this disease. Genetic reduction of Beclin 1 expression increased intraneuronal Aβ accumulation, extracellular Aβ deposition, and neurodegeneration [145]. Increasing Beclin 1 levels by lentiviral expression reduced both intracellular and extracellular amyloid pathology in these transgenic mice. This study suggests that decreased, not increased, autophagy promotes Alzheimer’s disease progression. Further, enhancing autophagy by increasing Beclin 1 levels may have therapeutic potential in this disease.
6.3. Autophagy and lysosomal storage diseases
Niemann–Pick type C is a neurodegenerative lipid storage disorder characterized by a disruption of sphingolipid and cholesterol trafficking caused by mutations in either of two genes, npc1 and npc2. The disease produces cognitive impairment, ataxia and death, often in childhood. Cells deficient in npc genes show increased expression of Beclin 1 and LC3-II, the autophagosome-specific form of LC3, suggesting autophagy is induced [146]. Increased levels of LC3-II have also been observed in npc-deficient brain tissue [147]. npc-Deficient cerebellar Purkinje neurons undergo a cell death that was proposed to depend on autophagy [148], suggesting increased autophagy may be harmful for neurons in NPC patients.
Most lysosomal storage diseases are caused by deficiencies of lysosomal hydrolases, leading to accumulation of undegraded substrate and other material in the lysosomal compartment. Lysosomal accumulation of substrates can also affect autophagosome–lysosome fusion. Autophagosomes accumulate in brain and isolated cell lines of mouse models of two lysosomal storage diseases associated with severe neurodegeneration, multiple sulfatase deficiency and mucopolysaccharidosis type IIIA [149]. Significantly reduced colocalization of the lysosomal membrane protein LAMP-1 with the autophagosome marker LC3 indicates that the fusion of lysosomal compartments with autophagosomes is impaired. In addition, cell lines isolated from these mice have decreased ability to degrade aggregate-prone proteins and show accumulation of polyubiquitinated proteins and non-functional mitochondria. Thus, neurodegeneration observed in many lysosomal storage diseases may be at least partially due to impaired autophagic degradation, which is particularly vital for neurons.
6.4. Autophagy and muscle disorders
Autophagic vacuoles are a frequent feature in numerous muscular disorders. Such a pathological situation can be observed in patients suffering from Danon disease, an inherited disease resulting from null mutations in the lysosomal membrane protein LAMP-2 [83]. LAMP-2 deficiency leads to a fatal cardiomyopathy and myopathy sometimes associated with mental retardation [150]. Accumulation of autophagic vacuoles in the heart and skeletal muscle are hallmarks of the disease [84]. Studies in LAMP-2 deficient mice revealed in part similar findings [85]. Fifty percent of these mice die at an early postnatal age with massive accumulation of autophagic vacuoles in several tissues including liver, pancreas, spleen, kidney, lymph nodes, neutrophilic leukocytes, skeletal muscle, and heart. Autophagic vacuoles containing single mitochondria were frequently observed in cardiomyocytes [151] indicating that mitochondria are a main target for autophagic degradation in muscle tissues. These cellular alterations lead to a reduced contractility and an increased size of the heart in LAMP-2 knockout mice. This is in agreement with the finding that cardiomyopathy is the hallmark in Danon disease patients [83]. Biochemical and electron microscopy studies reveal that a retarded consumption, rather than increased formation, of autophagic vacuoles leads to their accumulation [152]. LAMP-double deficient fibroblasts lack both LAMP-2 and the structurally related LAMP-1 protein. These cells show a defect in the final maturation steps of late autophagic vacuoles, involving retarded fusion with lysosomes [33], [119]. Interestingly, recruitment of the small GTPase Rab7 to autophagosomes is retarded in these cells [32].
Inhibition of lysosomal fusion using hydroxy-chloroquine causes similar vacuolar alterations and myopathies to Danon disease, confirming the important role of lysosome–autophagosome fusion for muscle cell physiology [153]. Impaired autophagosome maturation may also be related to other types of diseases such as X-linked myopathy with excessive autophagy, infantile autophagic vacuolar myopathy, adult-onset autophagic vacuolar myopathy with multiorgan involvement, and X-linked congenital autophagic vacuolar myopathy [154]. The molecular defects in these disorders are still unknown.
Although altered autophagy has been observed in various heart diseases, including cardiac hypertrophy and heart failure, it remains unclear whether autophagy plays a beneficial or detrimental role in these diseases. As mentioned earlier, tissue-specific deletion of atg5 in heart causes cardiac hypertrophy and contractile dysfunction [86]. In addition, increased levels of ubiquitinated proteins and abnormal mitochondria are found, especially after treatment with pressure overload or β-adrenergic stress. This suggests that autophagy is needed in the heart to ensure the availability of sufficient energy substrates and to control cardiomyocyte size and global cardiac structure and function.
6.5. Common aspects in autophagy and phagocytosis
As described above, autophagy plays a role in innate immunity against intracellular pathogens [104], [105] by clearing microbes directly via ingestion into autophagosomes for subsequent degradation in autolysosomes [108], [155]. Similar to the process of intracellular defense, phagocytosis is an evolutionary conserved mechanism involved in the removal of extracellular organisms. Interestingly, it was found that the phagocytic and autophagic pathways are linked. Toll like receptor (TLR) activation triggers the recruitment of autophagy proteins LC3, Atg5 and Atg7 to the phagosomal pathway. Before these events Beclin 1 and class III PI3-kinase activity are found in phagosomes [156]. These autophagy-specific proteins are recruited to the phagosome, while almost no classical autophagosomes are observed in the cells. Phagosome fusion with lysosomes is then initiated, leading to acidification and killing of the ingested organisms. Thus, engaging the autophagy pathway via TLR signaling (especially TLR7 through its binding to single-stranded RNA) enhances phagosome maturation and destruction of pathogens [157]. This underscores the intimate link between autophagy and phagocytosis.
As mentioned earlier, M. tuberculosis is able to survive inside macrophages by preventing the fusion of phagosomes with lysosomes, but induction of autophagy bypasses the maturation defect, leading to phagolysosome formation and bacterial killing [108]. Autophagy induction induces the localization of Beclin 1 and LC3 to phagosomes, suggesting the phagosomes are diverted to an autophagosome-like compartment that is then able to fuse with lysosomes.
The association between autophagy and phagocytosis is also underlined by studies with cells lacking either one or both LAMPs [33]. As described above, autophagosome–lysosome fusion is impaired in LAMP-double deficient cells [32]. Whereas macrophages and fibroblasts from LAMP-1 or LAMP-2 single-deficient mice display normal fusion of lysosomes with phagosomes, in LAMP-double knockout fibroblasts phagosomes are unable to recruit late endosomal/lysosomal markers and phagocytosis is arrested prior to the acquisition of Rab7 [158]. Interestingly, the maturation of Neisseria-containing phagosomes is also disturbed and cells lacking both LAMP proteins fail to kill the engulfed pathogens [159]. The maturation block caused by LAMP deficiency is at least partially due to the inability of autophagosomes and phagosomes to undergo dynein/dynactin-mediated centripetal movement along microtubules towards lysosomes [158]. Interestingly LAMP-2 single knockout mice show an impaired phagosomal maturation in neutrophilic leucocytes. The impairment of this innate immune defense mechanism leads to periodontitis, which is one of the most widespread infectious diseases worldwide. The retarded clearance of bacterial pathogens is due to an inefficient fusion between lysosomes and phagosomes, leading to less efficient killing of the ingested pathogens [160], [161]. Neutrophils of the LAMP-2 knockout mice also contain an accumulation of autophagic vacuoles [85], [160], which is likely also due to impaired fusion of autophagosomes with lysosomes.
Taken together these observations indicate that fusion with lysosomes is required to successfully complete both autophagosome and phagosome maturation that is necessary for efficient degradation of the cargo. Further, the results show that the maturation of autophagosomes and phagosomes share common features, because both processes are impaired with similar tissue and cell specificity in LAMP-2 deficient mice and LAMP-double deficient cell lines.
7. Conclusions
Degradation of cytosolic proteins in lysosomes via autophagy has turned out to have numerous, partly unexpected, roles in health and disease. Autophagy has been shown to contribute to innate and adaptive immunity and longevity, and to the prevention of cancer and neurodegeneration, just to mention a few of its newly-revealed functions. Treatments for human diseases that specifically target autophagy do not yet exist. It is likely, however, that such treatments will emerge in the future, once the molecular mechanisms of the processes involved in autophagy regulation have been clarified and suitable inducers and inhibitors for clinical trials have been identified.
Authors : Yong-Bo Hu Eric B Dammer Ru-Jing Ren Gang Wang
Abstract
The endosomal-lysosomal system is made up of a set of intracellular membranous compartments that dynamically interconvert, which is comprised of early endosomes, recycling endosomes, late endosomes, and the lysosome. In addition, autophagosomes execute autophagy, which delivers intracellular contents to the lysosome. Maturation of endosomes and/or autophagosomes into a lysosome creates an unique acidic environment within the cell for proteolysis and recycling of unneeded cellular components into usable amino acids and other biomolecular building blocks.
In the endocytic pathway, gradual maturation of endosomes into a lysosome and acidification of the late endosome are accompanied by vesicle trafficking, protein sorting and targeted degradation of some sorted cargo. Two opposing sorting systems are operating in these processes: the endosomal sorting complex required for transport (ESCRT) supports targeted degradation and the retromer supports retrograde retrieval of certain cargo.
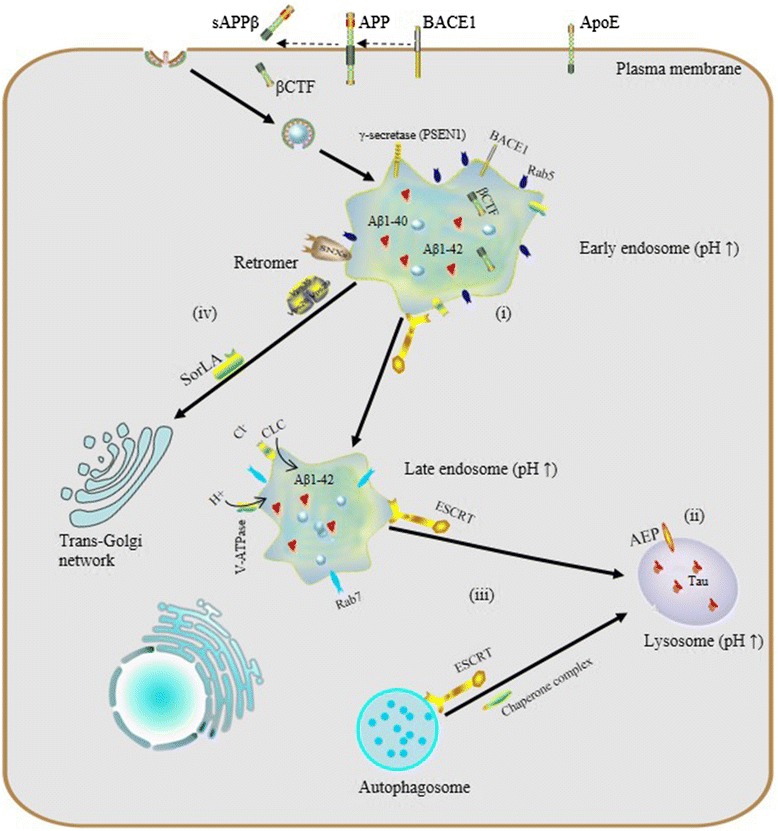
The endosomal-lysosomal system is emerging as a central player in a host of neurodegenerative diseases, demonstrating potential roles which are likely to be revealed in pathogenesis and for viable therapeutic strategies. Here we focus on the physiological process of endosomal-lysosomal maturation, acidification and sorting systems along the endocytic pathway, and further discuss relationships between abnormalities in the endosomal-lysosomal system and neurodegenerative diseases, especially Alzheimer’s disease (AD).
The endosomal-lysosomal system is a series of organelles in the endocytic pathway where various cargo molecules required for normal cellular function are internalized, recycled and modulated. Recently, mounting evidence has suggested that abnormalities in both endosomes and lysosomes, or dysregulation in their trafficking, play an important role directly in a surprising host of neurological dysfunctions, represented by AD, Parkinson’s disease (PD), and Lewy body dementia (LBD) [1–3]. Thus, the endosomal-lysosomal system is emerging as a key to understanding the mechanisms underlying both protein degradation and neurodegeneration. Here, we intend to summarize advances in the study of the endosomal-lysosomal system, with a focus on compartmentalized organization of trafficking routes, sorting machinery and their relationships to neurodegeneration.
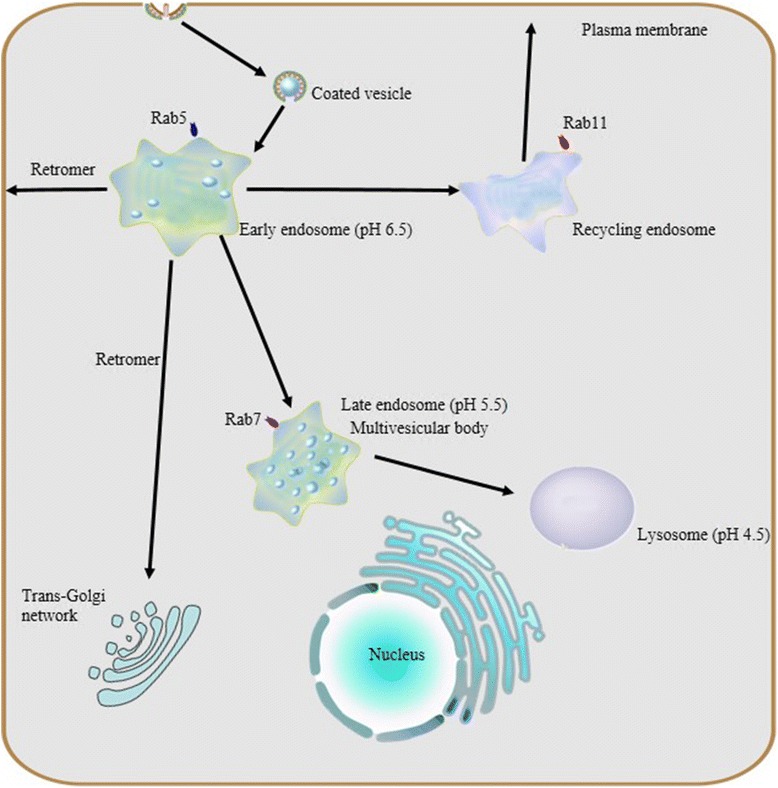
Endocytosis and endosome maturation. Protein internationalization is often dependent on a ubiquitous clathrin-mediated mechanism. Subsequent to internationalization, cargo proteins are transported to early endosomes via endocytic vesicles under the control of Rab5. Early endosomes serve as the major sorting stations where proteins can be sorted into recycling endosomes for recycling back to the cell surface, into a retrogradee pathway mediated by retromer to be sent back to the trans-golgi network (TGN), or into a degradation pathway for eventual targeting to the lysosome. As the number of intraluminal vesicles increases, early endosomes mature into late endosomes, and then late endosomes fuse with lysosomes.Due to their different capacities of acidification, a stable pH gradient is established in different compartments during the maturation process: early endosomes maintain pH at about 6.5, late endosomes at about 5.5 and lysosomes at about 4.5
A pH gradient established by vacuolar ATPase and chloride channels
Vacuolar ATP-dependent acidification
As mentioned above, the endosomal and lysosomal compartments share at least one similar significant characteristic: low intraluminal pH. These sealed acidic compartments provide an appropriate environment for optimal degradation of substrate cargo and recycling of their cognate receptors. As macromolecules are transported along the endosomal-lysosomal pathway, the internal pH of both endosomes and lysosomes decreases rapidly due to vacuolar acidification.
Previous research revealed that the same H+-ATPase, also known as vacuolar ATPase (V-ATPase), acidifies both endosomes and lysosomes. V-ATPase, differs from Na+,K+-ATPase in the plasma membrane, Ca2+-ATPase in the sarcoplasmic reticulum, and F1,F0-ATP synthase in mitochondria, in that it does not require a coupled influx of permeant anions [19]. Sulfhydryl alkylating reagents such as N-ethylmaleimide inhibit the V-ATPase dependent acidification of the endosomal-lysosomal system [19–21] as well as the specific inhibitor Bafilomycin A1 [22, 23].
V-ATPase is a unique class of ATPase present throughout the membranes which constrain the endocytic pathway, including the trans golgi network (TGN). V-ATPase, as a protein complex, is composed of two multimeric subunits, V1 in the cytoplasmic domain and V0 within the vacuolar membrane; the activity of V-ATPase depends on the dynamic assembly of these. V-ATPase is widely expressed in eukaryotic cells and serves as the master regulator of vesicular acidification in many subcellular membrane bound organelles. It also has important roles to play in vesicular trafficking and proteostasis.
Abnormalities and dysregulation of the endosomal-lysosomal system in neurodegeneration
Increasing attention being paid to the endosomal-lysosomal system has begun to elucidate a relationship between endosomal-lysosomal defects and neurodegeneration. In particular, robust pathology implicating endosomal-lysosomal disruption in AD has been well characterized. Here, we focus preferably on AD as a example of neurodegenerative disease and we believe that AD represents a general model of neurodegenerative diseases on abnormalities of the endosomal-lysosomal system occur along a continuum that includes early endosome changes, dysregulated acidification and sorting component defects.
The endosomal–lysosomal system and AD. (i) endosomal enlargement, Rab5 overexpression and Aß accumulation; (ii) dysregulated endosomal-lysosomal acidification, pH elevation and tau aggregation; (iii) dysfunctional ESCRT complexes, defective autophagy and accumulation of intraluminal ubiquitinated proteins; (iv) defects of retromer: reduction of Vps26, Vps35 and SorLA disrupts the trafficking and processing of APP
Dysregulated acidification, cellular indigestion?
Endosomal-lysosomal pH defects are an emerging theme in mechanisms underlying a number of neurodegenerative diseases. To date, results from experiments in vivo and in vitro have revealed the importance of proper vesicular pH balance and optimal acidification in transporting and degrading cargo via the endocytic pathway [48, 49]. For instance, Lee et al. reported that, in presenilin1 (PSEN1)-deleted blastocysts, defective lysosome acidification was observed with a substaintially elevated lysosomal PH of 5.4 and PSEN1 is essential for the transport of mature V0a1 subunites of V-ATP to lysosomes for their acidification and proteolysis [50].
Specifically, dysregulation of acidification and intracellular pH perturbation could influence the activity of enzymes in endomembrane compartments, resulting in impaired clearance of protein aggregates downstream of elevated endomembrane system pH, or conversely, due to decreased cytoplasmic pH. Regarding the latter, asparaginyl endopeptidase (AEP) is a typical pH-sensitive protein hydrolase the activity of which depends on the acidic pH of vesicular compartments. Predominantly localized in late endosomes, asparaginyl endopeptidase (AEP) specifically cleaves substrates with an asparagine residue at the P1 site. It is known that AEP can undergo reversible pH-dependent autoproteolytic activation, and in normal conditions, full-length pro-AEP is inactive [51].
As pH decreases from neutral to acidic, the activity of AEP gradually increases, such that it is partially activated at pH 4.5 and fully activated at pH 3.5, via removal of a cap that covers the active site. In AD patients, lysosomal acidification may be defective and it has been shown that the intracellular pH of neurons gradually decreases with aging [52] and more so with lactic acid elevation seen in AD cortex [53], so ectopic AEP activation or activity after leakage of active enzyme from late endosomes or lysosomes may be increased. AEP is involved in pathological tau degradation. Specifically, AEP generates tau fragments that form insoluble fibrils and result in neurotoxicity and neuropathological changes in AD [54, 55].
Increased endosome and lysosome pH is expected to have global effects on the proteome, particularly membrane proteins which rely on this pathway for their regulation and degradation. Interestingly, studies of microglia in culture have shown that in the absence of inflammatory IL-6 signaling, microglia do not achieve a sufficiently low lysosomal pH to degrade Aß, while after stimulation, CLC7 trafficking to lysosomes increases and pH drops sufficiently to improve Aß clearance [49, 56].
We have recently performed a systematic look at the proteomic effects of defective endosomal-lysosomal pH in a cellular model, in order to develop a better understanding of the global changes in the proteome that follow inhibition of V-ATPase and could be considered together as a signature or biomarker of defective vacuolar acidification [57, 58], which would be expected to have an overlap with changes seen in AD and/or other conditions which may be subject to this often age-dependent defect.
Indeed, blocking lysosomal degradation with bafilomycin A1 affects a significant increase in global K63 polyubiquitin linkages, which also occurs in AD, but AD brain global ubiquitin linkage profiling shows changes in other linkages as well [59]. Since K63 linked ubiquitin is not targeted to the proteasome, but does increase with V-ATPase acidification in the model of lysosomal insufficiency, the increase in K63 linkages seen in AD implicates accumulation of ubiquitinated proteins with obligate ESCRT-mediated degradation. Thus, trafficking, inflammatory signaling, and cell-type specific roles of dynamic lysosomal acidification are becoming increasingly appreciated for potential roles in AD pathogenesis.
Finally, it is important to point out that the general processes of endocytosis and endosomal-lysosomal dysregulations above-mentioned, have profoundly distinct implications for potential functions associated with other neurodegenerative diseases, such as PD, ALS, and Frontotemporal lobe degeneration (FTLD).
Conclusions and perspective
The endosomal-lysosomal system is a complex and highly dynamic process, where internalized transmembrane proteins, receptors, receptor ligands, and some soluble extracellular proteins are transported, sorted, and/or degraded. In recent years, particular attention has been paid to the endosomal-lysosomal system because it is involved in almost all of the neurodegenerative diseases, even though how it does so in each still remains unclear. Ongoing future studies will investigate both common and cell-type (or even local membrane region) specific trafficking and proteostasis pathways involving the endosomal-lysosomal system as well as the larger endomembrane system.
For example, a better understanding of distinct roles that ubiquitination plays in ESCRT-mediated proteostasis (and even lipid droplet homeostasis [79] which appears to be dysregulated in glia in neurodegeneration [47, 80]) could help to predict and ultimately therapeutically address the onset and progression of neurodegenerative diseases for specific individuals or sub-populations. This milieu of membrane-bound proteins that dynamically sorts cargo enriched for signaling, inflammation, and neurotrophic functions—among others—promises to provide a mother lode of new therapeutic targets for amelioriating neurodegenerative diseases, but the exploration also promises to be challenging, requiring the development of novel techniques and insight.
pH-Dependent Entry of Severe Acute Respiratory Syndrome Coronavirus Is Mediated by the Spike Glycoprotein and Enhanced by Dendritic Cell Transfer through DC-SIGN
June 2004
ABSTRACT
The severe acute respiratory syndrome coronavirus (SARS-CoV) synthesizes several putative viral envelope proteins, including the spike (S), membrane (M), and small envelope (E) glycoproteins. Although these proteins likely are essential for viral replication, their specific roles in SARS-CoV entry have not been defined. In this report, we show that the SARS-CoV S glycoprotein mediates viral entry through pH-dependent endocytosis. Further, we define its cellular tropism and demonstrate that virus transmission occurs through cell-mediated transfer by dendritic cells. The S glycoprotein was used successfully to pseudotype replication-defective retroviral and lentiviral vectors that readily infected Vero cells as well as primary pulmonary and renal epithelial cells from human, nonhuman primate, and, to a lesser extent, feline species. The tropism of this reporter virus was similar to that of wild-type, replication-competent SARS-CoV, and binding of purified S to susceptible target cells was demonstrated by flow cytometry. Although myeloid dendritic cells were able to interact with S and to bind virus, these cells could not be infected by SARS-CoV. However, these cells were able to transfer the virus to susceptible target cells through a synapse-like structure. Both cell-mediated infection and direct infection were inhibited by anti-S antisera, indicating that strategies directed toward this gene product are likely to confer a therapeutic benefit for antiviral drugs or the development of a SARS vaccine.
The severe acute respiratory syndrome coronavirus (SARS-CoV) is the likely cause of an acute infectious respiratory disorder identified in highly lethal outbreaks during the past year (10, 18, 21, 32, 40). Infection is characterized by acute flu-like symptoms that progress to a severe febrile respiratory illness with significant mortality. Coronaviruses, comprising a genus of the Coronaviridae family, are enveloped positive-strand RNA viruses. In general, coronaviruses cause respiratory and enteric diseases in humans and domestic animals (15, 20). Two previously known human coronaviruses caused only mild upper respiratory infections (15, 20). In contrast, a highly pathogenic, severe respiratory disease is caused by the SARS-CoV, especially in the elderly (44). Coronaviruses can be divided into three serologically distinct groups (15). Phylogenetically, SARS-CoV is not closely related to any of the three groups (26), though it is most similar to the group II coronaviruses (33, 36).
Although the organization of the SARS-CoV genome is related to that of animal coronaviruses, its genetic sequence is unique, and the structure and function of its gene products are not known. At least 14 open reading frames (ORFs) can be identified in its genome (26, 34, 36). Among these, the replicase/transcriptase genes are located in the 5′ portion of the genome. At its 3′ end, the four major structural proteins (S, M, N, and E) are made through different subgenomic RNAs. Based on comparison to animal coronaviruses, three structural gene products are predicted to be present on the viral envelope: the spike (S), membrane (M), and small envelope (E) proteins (20, 26, 34). The structure of the SARS-CoV envelope differs in some respects from that of other enveloped viruses, such as retroviruses and lentiviruses, many of which contain one viral envelope protein.
Envelope or spike proteins from enveloped viruses have been used to pseudotype retroviral and lentiviral vectors for functional and gene transfer studies (29, 35, 43, 45); however, whether coronavirus glycoproteins could pseudotype these viruses was unknown. Here we report that replication-defective retroviral (Moloney murine leukemia virus) and lentiviral (human immunodeficiency virus type 1 [HIV-1]) vectors can be pseudotyped with the SARS-CoV S protein, and the properties of S related to entry have been defined. Using these pseudoviruses, we were able to determine the relative contributions of SARS-CoV envelope proteins to viral entry and fusion and to examine the roles of these different viral envelope gene products with respect to entry, cell specificity, and potential inhibition of viral replication.
Pertinent Extract:
In contrast, influenza and Ebola viruses are prototypes for viruses that utilize a pH-dependent endocytotic pathway (43). To determine the pathway utilized by the SARS-CoV, the pH dependence of the SARS-CoV S-pseudotyped lentiviral vector was analyzed. Addition of ammonium chloride, which prevents acidification of the endosome, caused a dose-dependent reduction in viral entry (Fig. (Fig.1B,1B, left) at concentrations similar to those described for other pH-dependent viral glycoproteins (3, 11, 43). This effect was also observed with another inhibitor of endosomal acidification, bafilomycin, also in a dose-dependent fashion (Fig. (Fig.1B,1B, right).
Previous research from Meridian Institute Article :
Possible Relevance to SARS
The World Health Organization has concluded that SARS is produced by a new virulent strain of coronavirus. Specific research on the possible pH dependency of the SARS virus has not yet been done. It is well known that coronavirus infectivity is exquisitely sensitive to pH. For example, the MHV-A59 strain of coronavirus is quite stable at pH 6.0 (acidic) but becomes rapidly and irreversibly inactivated by brief treatment at pH 8.0 (alkaline). Human coronavirus strain 229E is maximally infective at pH 6.0. Infection of cells by murine coronavirus A59 at pH 6.0 (acidic) rather than pH 7.0 (neutral) yields a tenfold increase in the infectivity of the virus.
ChooseLife : If the strain of coronavirus responsible for SARS shares the pH characteristics of these other coronaviruses that are pH-dependent, this could be a valuable clue to effective prevention and treatment strategies for this potential epidemic. Perhaps keeping a balanced or slightly alkaline pH environment for the body’s tissues can provide viral protection or enhanced healing for SARS and common viral agents that cause respiratory infections.
Inter-related to this, is research on MUC5B, which has shown that those of lower pH, are much more prone to having inhibited mucous membrane formation:
“Moreover, we demonstrate that the conformation of these highly entangled linear polymers is sensitive to calcium concentration and changes in pH. In the presence of calcium (Ca2+, 10 mM) at pH 5.0, MUC5B adopted a compact conformation which was lost either upon removal of calcium with EGTA, or by increasing the pH to 7.4. These results suggest a pathway of mucin collapse to enable intracellular packaging and mechanisms driving mucin expansion following secretion. They also point to the importance of the tight control of calcium and pHduring different stages of mucin biosynthesis and secretion, and in the generation of correct mucus barrier properties. “
ChooseLife Related Thoughts :
The above shows that there are multiple potential protective methodologies in play, some people may feel a glass of cold water with 1/2 teaspoon of Sodium Bicarbonate every two hours on the first day may be effective (outlined at the bottom of this page), this is one method I would consider myself (Arm and Hammer or Bobs Mill being Aluminium free). Also small Sips of highly Alkaline Milk of Magnesia, every hour, may coat the upper respiratory regions fairly well and rapidly bring up the pH, out of the greater danger zones of lower pH < 6.5 (this is my go to for my kids with sniffles or worse), I would likely do this myself for this situation.
Personally I am going to use this outbreak as a good time to bring my own (and childrens) pH up, using methods as above, plus make some Moreless Alkalising Mineral Mixture, which both Alkalises and significantly raises the Calcium levels in the body but in a complexed form (pre-bonded to Molasses or Honey) which does not hamper the Mucous membrane process outlined above, which as shown above in the scientfic literature is exactly what our bodies need to be ready to either repel, or minimise the effects of such threats.
“The proven value of Bicarbonate of Soda as a therapeutic agent (from a letter to the Church and Dwight Company):
In 1918 and 1919 while fighting the Flu with the U.S. Public Health Service it was brought to my attention that rarely any one who had been thoroughly alkalinized with bicarbonate of soda contracted the disease, and those who did contract it, if alkalinized early, would invariably have mild attacks. I have since that time treated all cases of Cold, Influenza and LaGripe by first giving generous doses of Bicarbonate of Soda, and in many, many instances within 36 hours the symptoms would have entirely abated.
Further, within my own household, before Women’s Clubs and Parent-Teachers’ Association, I have advocated the use of soda as a preventative for ‘Colds’, with the result that now many reports are coming in stating that those who took ‘Soda’ were not affected, while nearly everyone around them had the ‘Flu’.
…An occasional three-day course of the Bicarbonate of Soda increases the alkalinity of the blood, assists elimination and increases the resisting power of the body to all Infectious Diseases…
Whenever taking a bicarbonate solution internally, the soda should be dissolved in cold water. In the event of a threatened attack we recommend the following treatment: During the first day take six doses of half a teaspoon of Bicarbonate of Soda in a glass of cool water, at about two hour intervals.”
The investigation was performed using small, bacteria-eating organisms called Caenorhabditis elegans, here imaged using Oregon Green-labeled dextran and 748 laser-scanning confocal microscopy.
When food we’ve swallowed reaches our stomachs, it finds an acidic environment. The low pH in the stomach helps to begin digestion—and has been thought to kill the bacteria that hides in food that otherwise could harm our bodies.
However, recent work from the Ackley and Chandler labs in the Department of Molecular Biosciences at the University of Kansas runs counter to this idea, instead suggesting lower pH in the digestive tract may make some bacterial pathogens even more harmful.
Their findings, published in the peer-reviewed journal PLOS Pathogens, could have implications for addressing the crisis of antibiotic resistance in bacterial infections around the world.
The investigation was performed using small, bacteria-eating organisms called Caenorhabditis elegans.
“These wormlike animals are transparent, so we can watch things that happen inside them quite easily,” said co-author Brian Ackley, associate professor of molecular biosciences at KU. “Using pH-sensitive chemicals developed at KU, called Kansas Reds, we were able to monitor the pH inside the digestive system and watch what happens when they eat harmful bacteria, compared to nonharmful bacteria.”
According to the KU researchers, under normal conditions while feeding on healthy bacteria, C. elegans digestive tracts are moderately acidic compared to human stomachs. But these model species’ stomachs also show regional differences within the digestive tract. When they ingest pathogens, they neutralize the acidic environment.
This observation suggested the animals could discriminate between good and bad bacteria, and harmful bacteria prompted a less acidic digestive tract in C. elegans—a result that runs counter to what one might expect if the acidic environment was generated to kill bacteria.
To test this, the researchers used animals with mutations in genes that helped regulate the pH in their digestive tracts.
“When animals had a more acidic digestive system, they were more likely to be affected by pathogenic bacteria—again counter to what one might guess if acidity was useful in killing harmful bugs that might sneak into the body with food,” Ackley said. “Our lab teams were able to show the effect on the animals was specifically due to the pH by adding a base to buffer the digestive tract. We used bicarbonate, the same agent our bodies use to neutralize stomach contents when they pass into our intestines. Neutralizing the pH in the mutant animals reverted the accelerated infection by the pathogenic bacteria.”
The KU researcher said different species react differently when their bodies sense pathogenic bacteria—but some biological reactions are common to many animals.
“A general response involves the creation of chemicals, like hydrogen peroxide or hypochlorous acid—aka bleach—near the bacteria, and then having specialized immune cells eat the dying bacteria,” Ackley said. “To keep our bodies safe, the immune system only deploys these defenses when it’s sure it is being invaded. The work in C. elegans may suggest a way the body can have these defenses ready to go at a moment’s notice—that is, keep the chemical environment in a moderately acidic state where making those chemicals is difficult, then, upon infection, simply neutralize the environment to deploy the defenses.”
Ackley’s KU colleagues on the work were lead author Saida Benomar, Patrick Lansdon and Josephine R. Chandler of the Department of Molecular Biosciences, along with Aaron Bender of the Department of Medicinal Chemistry, and Blake R. Peterson of The Ohio State University.
The researchers believe there may be reasons to believe these systems could work similarly in people.
The genes they studied in C. elegans also exist in humans and control parts of the immune system. Further, research in other labs has shown occasions in humans where problems with regulating pH are associated with increased risk of infection. Moving forward, the researchers want to understand the mechanism at a deeper level.
“Our goal is to boost this natural defense system in people as a way to either avoid or reduce the use of antibiotics,” Ackley said. “Right now, our antibiotic use is unsustainable, and bacteria are evolving resistance at an alarming rate. If the system discovered in C. elegans is in fact still present in humans, it would suggest bacteria are much slower to adapt to this defensive strategy than they are to antibiotics.”
ChooseLife : Carey Reams stated that the production and regulation of stomach acid, is firstly regulated by the Liver, and, that primarily it’s production requires Calcium Salt in the form of Calcium Gluconate. Reams often proposed that a diet lower in Protein was better for health, so this may align to this concept, that those eating more meat or plant proteins may have higher stomach acid production, and, hence a generally lower pH. Then becoming more susceptible to these type of infections?